













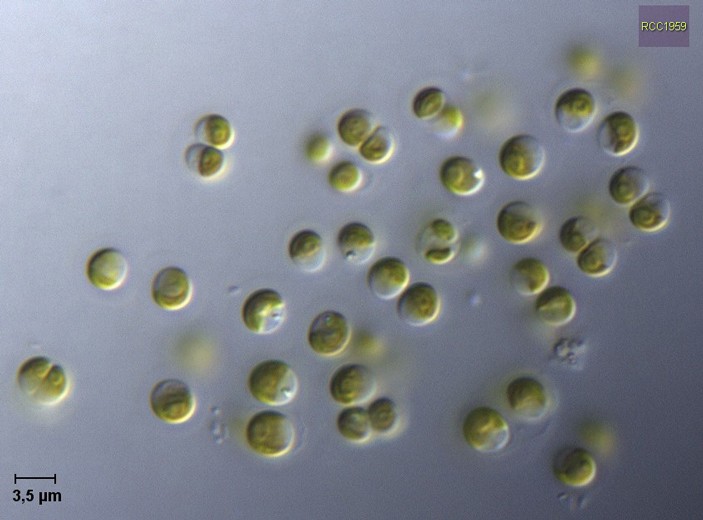


Рис. 1. Prasinoderma: одиночные безжгутиковые клетки правильной формы, с ядрами и хлоропластами внутри. Видно, что некоторые клетки прямо в момент фотографирования находятся на разных стадиях деления — на это время они, конечно, одиночными быть перестают. Длина масштабного отрезка — 3,5 микрона. Источник фото: http://roscoff-culture-collection.org/rcc-strain-details/1959
Прочтение ядерного генома микроскопической океанской водоросли празинодермы привело ученых к выводу, что эта водоросль принадлежит к особой эволюционной ветви. Раньше считалось, что группа зеленых растений (Viridiplantae) состоит всего из двух больших ветвей: хлорофит («зеленых водорослей») и стрептофит, которые раньше назывались харофитами и к которым относятся наземные растения. Но празинодерма, похоже, принадлежит к третьей ветви, идущей от самого корня древа зеленых растений.
Ботаника — одна из самых старых биологических дисциплин. Ее основателем считается Теофраст, ученик Аристотеля, второй глава школы перипатетиков. Ясное дело, что за две с лишним тысячи лет она накопила огромный методологический опыт (см., например: Г. Ю. Любарский, 2015. Рождение науки). Но интересно, что едва ли не всё это время оставался нерешенным вопрос о границах ботаники, о том, какие организмы являются растениями, а какие нет. В «пограничную ситуацию» попадали не только грибы, которых французский ботаник Себастьян Вайан (Sébastien Vaillant) в начале XVIII века обозвал по этой причине порождениями дьявола. Ничуть не меньшей проблемой были всевозможные водоросли: зеленые, красные, бурые, золотистые и другие.
Изучение водорослей долгое время тормозилось трудностью детальных исследований строения клеток. Но когда такие исследования начались, они довольно быстро привели ботаников к мысли «о возможности происхождения комплекса организмов, именуемого растениями, не от одного общего корня, а разными путями» (А. А. Уранов, 1979. Методологические основы систематики растений (в их историческом развитии)).
И в самом деле, допустим, что «растения» — это автотрофные эукариоты. Мир автотрофных эукариот разнообразен чрезвычайно. Еще в первой трети XX века знаменитый ботаник Адольф Пашер (Adolf Alois Pascher) в серии работ обосновал идею, что мир «растений» — это несколько эволюционных стволов, развивавшихся параллельно, но независимо друг от друга. Каждый из них начинался с одноклеточных жгутиковых форм.
Эта идея оказалась совершенно верной. В конце XX века, в эпоху великого пересмотра системы живой природы, английский биолог Томас Кавалье-Смит (Thomas Cavalier-Smith) формально выделил группу под названием Viridiplantae, что значит «зеленые растения». Viridiplantae — это эволюционный ствол растений в наиболее привычном нам смысле. К ним относятся высшие растения — споровые, голосеменные, цветковые — и многочисленные родственные им водоросли, большинству из которых свойствен зеленый цвет. Ближайшие «внешние» родственники Viridiplantae — красные водоросли и глаукофитовые водоросли, вместе с которыми они образуют супергруппу Archaeplastida. А вот бурые, золотистые, диатомовые или эвгленовые водоросли относятся вообще к другим супергруппам эукариот (см. рис. 2 в статье: Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019).
Что такое супергруппа? В XXI веке этим словом стали для удобства называть крупнейшие эволюционные стволы эукариот: такие, которые можно пересчитать по пальцам если не одной руки, то уж хотя бы обеих рук. В терминах классической ранговой систематики супергруппа — это, пожалуй, надцарство. Тогда Viridiplantae — это царство. Именно такой ранг и дал этой группе Кавалье-Смит при ее выделении (T. Cavalier-Smith, 1981. Eukaryote kingdoms: seven or nine?). Так что понятие «царство Viridiplantae» можно употреблять вполне законно, хотя, конечно, надо учитывать, что в современной биологической системе все подобные ранги до некоторой степени условны.
Разнообразие зеленых растений колоссально. Какое-то время назад можно было считать, что именно к царству Viridiplantae относятся и самые крупные, и самые мелкие эукариоты. Мельчайшее зеленое растение — одноклеточная морская водоросль Ostreococcus, средний диаметр которой 0,8 микрона. А крупнейшее зеленое растение — гигантская секвойя (Sequoiadendron giganteum), которая достигает высоты около 95 метров и способна претендовать на звание самого большого живого организма на Земле. К сожалению, сейчас ее рекорд побит: самым большим организмом считается не растение, а гриб. Это мицелий одного из видов рода опёнок (Armillaria ostoyae), который растет на западе Северной Америки, в Орегоне, и покрывает площадь больше девяти квадратных километров. Но среди организмов, обладающих истинной многоклеточностью и тканевой организацией, секвойя остается рекордсменом. В любом случае эти примеры показывают, как широки эволюционные возможности зеленых растений. На Земле очень мало сообществ, в которых бы они не встречались.
Эволюционное древо Viridiplantae устроено довольно сложно (см. Сценарий происхождения высших растений во многом похож на сценарий происхождения животных, «Элементы», 30.04.2020). До недавнего времени считалось бесспорным, что оно распадается на два эволюционных ствола: стрептофиты (Streptophyta) и хлорофиты. В обоих стволах есть и одноклеточные, и многоклеточные формы. Хлорофиты в целом разнообразнее стрептофит, но зато именно к стрептофитам относятся высшие (то есть наземные) растения.
Как выглядел общий предок стрептофит и хлорофит? Ну, наверняка это был одноклеточный жгутиконосец. Чтобы узнать про него больше, было бы неплохо найти эволюционную ветвь, происходящую непосредственно от него. Иными словами, нужна такая водоросль, которая бы входила в царство Viridiplantae, но не относилась ни к хлорофитам, ни к стрептофитам. Это и будет самая базальная — то есть, проще говоря, самая древняя — ветвь зеленых растений.
До сих пор все кандидаты на роль такой ветви «не выдерживали экзамена»: молекулярная филогенетика в конце концов всё же относила их к одной из двух больших подгрупп (например, мезостигмовые водоросли, которые оказались-таки базальными стрептофитами; см. Сценарий происхождения высших растений во многом похож на сценарий происхождения животных, «Элементы», 30.04.2020). В лучшем случае положение таких кандидатов оставалось спорным.
Но вот сейчас международная группа китайских, датских, бельгийских, канадских и южноафриканских исследователей сообщает, что удалось полностью прочитать ядерный геном одноклеточной морской водоросли, которая называется Prasinoderma coloniale (рис. 1, 2). Эта водоросль относится к пикоэукариотам — очень мелким одноклеточным эукариотам, размеры которых сравнимы с бактериальными (упомянутый выше остреококкус, представитель хлорофит, тоже к ним относится). Пикоэукариот много в океанском планктоне. Геном у празинодермы, как и следовало ожидать, оказался маленьким — всего 25,3 мегабазы (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Число белок-кодирующих генов у нее тоже невелико: чуть больше 7 тысяч. Полученный материал позволил провести тщательный молекулярно-филогенетический анализ — проще говоря, анализ родственных связей. И оказалось, что празинодерма, вероятно, образует сестринскую группу по отношению ко всем остальным Viridiplantae. Если это подтвердится — значит, искомая базальная ветвь найдена.

Рис. 2. Физическая карта ядерного генома Prasinoderma coloniale. Внешний круг — 22 хромосомы, внутренние круги отображают различные характеристики генома этих хромосом (например, долю ГЦ-пар). Иллюстрация из обсуждаемой статьи в Nature Ecology & Evolution
Тем самым в царстве Viridiplantae приходится выделять не две главные ветви, а три: Chlorophyta, Streptophyta и Prasinodermophyta (рис. 3). К последней на данный момент достоверно относится всего два рода, Prasinoderma и близкий к ней Prasinococcus. В классической ботанической систематике эти ветви, видимо, считались бы отделами. В обсуждаемой статье празинодермофиты описаны именно как новый отдел. Но современная систематика — филогенетическая (то есть основанная строго на родстве), поэтому в ней дело обстоит чуть сложнее: ветвь Prasinodermophyta должна получить одинаковый ранг с ветвью, включающей хлорофит и стрептофит вместе. А вот ранги самих хлорофит и стрептофит должны быть на ступеньку ниже: это следующий уровень ветвления древа. Проще говоря, если празинодермофиты — отдел (divisio), то хлорофиты и стрептофиты — подотделы (subdivisio). А если хлорофиты и стрептофиты останутся отделами, то празинодермофиты по логике вещей должны стать надотделом (superdivisio). Как ботаники распутают все эти номенклатурные хитросплетения, мы пока не знаем.

Рис. 3. Предполагаемое место празинодермофит в эволюции растений. Внизу — эволюционное древо, полученное авторами обсуждаемой работы. Следует заметить, что красные водоросли тут однозначно занимают более базальное положение, чем глаукофитовые. На многих других молекулярных деревьях более базальными оказывались, наоборот, глаукофиты; иногда эту проблему честно оставляют нерешенной (см., например: F. Burky et al., 2020. The new tree of eukaryotes). Как оно на самом деле, покажет будущее. Вверху — символическое изображение самих «героев» статьи с указанием их молекулярно-биологических характеристик: дополнительные белки светособирающего комплекса (Light harvesting complex, LHT), горизонтальный перенос генов от бактерий (Horizontal gene transfer, HGT), утрата способности к синтезу витаминов группы B, которые тоже приходится получать от бактерий, и синтез НАД через триптофан (показан его побочный продукт — пиколиновая кислота). Иллюстрация из статьи: M. Melkonian, H. Liu, 2020. A third phylum of green plants
Интересно, что представители древнейшей ветви зеленых растений оказались довольно-таки специализированными существами. Празинодерма обладает типичным коккоидным строением: одноклеточная, безжгутиковая, компактная, одетая в жесткую оболочку. Диаметр клеток — от 2 до 5 микрон (T. Hasegawa et al., 1996. Prasinoderma coloniale gen. et sp. nov., a new pelagic coccoid prasinophyte from the western Pacific Ocean). Отсутствие жгутиков означает, что у нее нет самостоятельной подвижности: клетки коккоидных водорослей могут только переноситься течениями воды. Океанские популяции таких водорослей существуют за счет быстрого размножения: какая-то часть клеток безвозвратно тонет, и новые клетки должны успевать приходить им на смену. Отсюда и давление отбора в сторону маленького размера генома: хорошо известно, что чем меньше геном, тем быстрее клетка делится (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015).
Неудивительно, что эволюционная линия празинодермы «разгружает» геном за счет всего, от чего можно отказаться. Например, она утратила несколько регуляторных генов (факторов транскрипции), которые, несомненно, были унаследованы зелеными растениями от предков: эти гены есть у хлорофит, у стрептофит, у красных водорослей и у глаукофит. Потеряла она и множество генов, продукты которых обычно служат для постройки растительных клеточных стенок, и гены, позволяющие синтезировать некоторые витамины (B1, B7 и B12). Эти витамины празинодерма, судя по всему, получает от бактерий, тоже живущих в океанском планктоне.
А вот гены, обеспечивающие постройку жгутиков (их довольно много — несколько десятков), сохранились у празинодермы почти в полном наборе, несмотря на то, что никаких жгутиковых стадий жизненного цикла у нее не обнаружено. Иными словами, безжгутиковость празинодермы еще не стала необратимой. Возможно, это означает, что уход этой линии водорослей в пикоэукариотную экологическую нишу произошел эволюционно недавно. Не исключается даже, что в ее жизненном цикле всё же есть жгутиковые стадии, просто пока не найденные.
Конечно, у празинодермы есть и собственные эволюционные приобретения. В первую очередь это кое-какие изменения в механизме фотосинтеза, связанные, возможно, с жизнью в относительно глубоких слоях воды — там мало света (исследованные экземпляры празинодермы были собраны в Северной Атлантике на глубине 150 метров). У празинодермы увеличено число генов, кодирующих белки светособирающих компексов: это означает, что в ее эволюционной линии произошло несколько уникальных генных дупликаций. Другие белки фотосинтеза у нее тоже имеют свои особенности.
Интересная «интрига» касается синтеза кофермента, который называется никотинамидадениндинуклеотид, сокращенно НАД. Это абсолютно необходимая молекула, без которой просто-напросто невозможен, например, процесс дыхания. У эукариот НАД может синтезироваться двумя разными путями: через аспартат или через триптофан (последний путь часто называют кинурениновым, по названию промежуточного продукта). У человека, например, НАД синтезируется через триптофан. У красных водорослей — тоже через триптофан, ферментов, позволяющих синтезировать НАД через аспартат, у них просто нет. А вот у остальных зеленых растений — и у хлорофит, и у стрептофит, включая наземные растения, и даже у глаукофит — НАД синтезируется только через аспартат.
Что касается празинодермы, то анализ ее генома показал неожиданную вещь. Празинодермофиты — единственные известные эукариоты, у которых есть оба пути синтеза НАД одновременно.
Объяснение этому дают следующее. Примитивным для эукариот является кинурениновый (иными словами, триптофановый) путь синтеза НАД. Однако общий предок супергруппы Archaeplastida получил два гена, позволяющих синтезировать НАД через аспартат, «в подарок» от цианобактерии, которую он поглотил и сделал первичным хлоропластом (C. M. Ternes, G. Schönknecht, 2014. Gene transfers shaped the evolution of de novo NAD+ biosynthesis in eukaryotes; об эволюции хлоропластов см. также: Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019). Ничего удивительного в этом нет, перенос генов из хлоропластов в ядро хозяйской клетки — дело обыкновенное. В дальнейшем и глаукофиты, и хлорофиты, и стрептофиты потеряли кинурениновый путь вообще, сохранив только аспартатный. А вот у празинодермофитов сохранился и тот, и другой. Это — крайне архаичное для зеленых растений состояние.
Справедливости ради надо сказать, что есть факты, которые в этот сценарий вписываются плохо. Во-первых, неясно, почему красные водоросли сохранили кинурениновый путь, но потеряли аспартатный (если он у них вообще когда-нибудь был). Во-вторых, гены аспартатного синтеза НАД имеют цианобактериальное происхождение только у глаукофит — одной из самых древних и примитивных ветвей архепластид. У зеленых растений, включая и празинодерму, эти гены тоже есть, и там они тоже явно заимствованы путем горизонтального переноса… но не от цианобактерий, а от других групп бактерий, не имеющих к хлоропластам никакого отношения. Похоже, они приобретались эукариотами несколько раз независимо.
В целом вопрос о том, насколько полно празинодермофиты отображают облик общего предка зеленых растений, остается открытым. Например, у этого общего предка наверняка были жгутики (как у упоминавшихся выше мезостигмовых водорослей, см. Mesostigma), а вот празинодерма жгутики потеряла. Да и геном у нее маловат. На архаичные черты тут, как всегда и бывает, наслоилась специализация — в данном случае в сторону превращения в пикопланктон, которому присущ четкий комплекс признаков: крайне мелкий размер, утрата самостоятельной подвижности, маленький геном, позволяющий быстро размножаться. Что ж, склонность к такой специализации явно возникала в эволюции эукариот не однажды. Нельзя исключить, например, что через такую эволюционную стадию когда-то прошли красные водоросли: именно этим можно было бы объяснить катастрофическое «обнищание» их генома (см. Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019). В общем, полученные ответы, как обычно, порождают новые вопросы, и это прекрасно.
Источник: elementy.ru





