













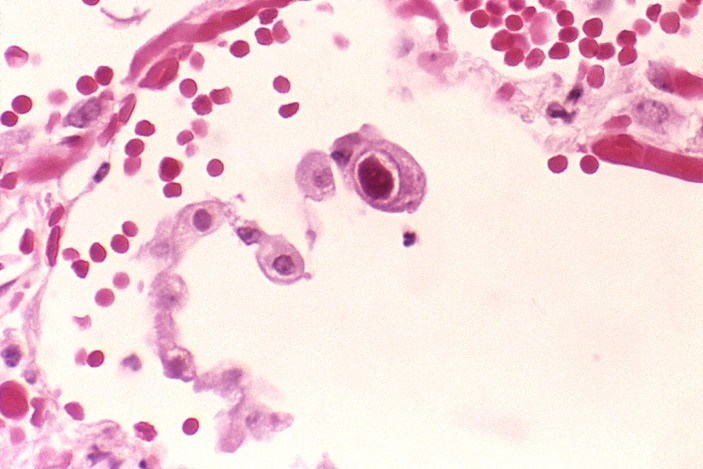

Рис. 1. Клетка легочного эпителия (альвеолоцит), пораженная цитомегаловирусом (в центре). Оправдывая свое название, вирус вызывает увеличение клеточного ядра. Фото с сайта ru.wikipedia.org
Рыночные отношения и естественный отбор имеют немало общих черт, а изучающие их науки — экономика и эволюционная биология — нередко заимствуют друг у друга идеи и концепции. Не зря Чарльз Дарвин вдохновлялся трудами экономиста Т. Роберта Мальтуса; концепция «Трагедии общин», описывающая противоречия между общими и частными интересами, во многом сформировала как современную экономическую теорию, так и понятие «эгоистичности» в представлениях современных эволюционистов, а математический аппарат теории игр в равной степени хорошо применим и для описания действий игроков на рынке, и для объяснения поведения животных. Еще один пример параллелизма между биологическими и экономическими концепциями, о котором пойдет речь в этой статье, — хеджирование рисков: попытка подстраховаться на случай возможных катастрофических событий на рынке / в естественной среде, пожертвовав некоторой частью экономического / эволюционного успеха. Хеджирование рисков в живой природе биологи называют «бет-хеджингом» — термином, заимствованным из лексикона игроков на тотализаторе. Бет-хеджинг заключается в стремлении уменьшить вариабельность своего репродуктивного успеха (например, избежать больших перепадов в числе выживших потомков) пусть даже за счет частичного уменьшения самого репродуктивного успеха. В каком-то смысле, бет-хеджинг — это стратегия «синицы в руке»: пусть потомков будет меньше, но зато их число будет более предсказуемым, а сами они будут приспособленными к более широкому спектру условий. В последнее время примеры подобной стратегии находят у самых разных живых организмов. В недавнем исследовании группа американских биологов изучала бет-хеджинг на примере одного из самых распространенных вирусов человека — цитомегаловируса. В ходе работы им удалось установить механизм, скрывающийся за более и менее агрессивными стратегиями эксплуатации хозяина, а также продемонстрировать, какую пользу вирусу может приносить каждая из них.
Что день грядущий мне готовит?
Мир непредсказуем. Эту простую истину в той или иной степени испытал на себе каждый, но есть те, для кого неопределенность будущего представляет собой каждодневную головную боль: это биржевые игроки, фанаты игры на тотализаторе, не объяснившиеся влюбленные и все живые существа, планирующие оставить потомство. Родители всегда немного похожи на генералов, хорошо готовых к уже прошедшим войнам, поэтому неудивительно, что, отпуская потомство в большой мир, они оказываются не в состоянии предсказать ожидающие его опасности. Да что мы — даже наши гены бессильны в этом вопросе…
В своей книге «Расширенный фенотип» (глава 3) известный эволюционист Р. Докинз замечает, что есть ряд фундаментальных причин, по которым естественный отбор не может быть абсолютно эффективен, а выработанные им адаптации всегда несовершенны, и первая из них — неспособность отбора идти в ногу со временем. Действительно, гены любого современного организма отбирались на протяжении долгих предшествующих эпох, когда условия окружающей среды были иными. Впрочем, не обязательно ходить так далеко. Возьмем, например, родителей (или родителя!) любого живого существа. По определению они были достаточно хорошо приспособлены к окружающей среде, чтобы оставить потомство. Наверняка и их потомки будут неплохо приспособлены к похожим условиям. Как правило, условия меняются не очень сильно, поэтому система работает неплохо. Более того, родители могут «подрегулировать» геном детей так, чтобы он максимально хорошо работал в обстоятельствах, в которых жили родители.
Типичным примером такой регуляции служит материнский эффект, когда родители «глушат» или, наоборот, усиливают экспрессию определенных генов потомства в соответствии с собственным жизненным опытом ( «Элементы» уже писали о материнском эффекте: Пищевое поведение у пещерной рыбы наследуется эпигенетически, 27.09.2012 и Неадаптивная фенотипическая пластичность затрудняет видообразование, 09.12.2015). Скажем, если самка коловратки — мелкого планктонного животного — живет в водоеме, где много хищников, то она «настраивает» гены своих дочерей (коловратки обычно размножаются партеногенетически) так, чтобы дочерние особи обладали длинными шипами, способными защитить от нападения хищника (J. J. Gilbert, 2016. Non-genetic polymorphisms in rotifers: environmental and endogenous controls, development, and features for predictable or unpredictable environments). Если же мать прожила свою жизнь в относительной безопасности, то и дочерям нет смысла обзаводиться дорогостоящей и бесполезной в отсутствие хищника защитой, поэтому они вырастают короткошипыми. И это при сходном генотипе! Подобная логика: в каких условиях родился, в тех и пригодился, или «Торгуй по-моему, так будешь не внакладе» — так в лавке говорил племяннику Купец», — краеугольный камень естественного отбора и… жизненный принцип многих людей. Поэтому настоящий кошмар для любых родителей — это возможные серьезные и плохо предсказуемые изменения условий окружающей среды. К жизни, похожей на родительскую, они детей приготовят, но что будет, если жизнь будет совсем иной? Что если желанная стабильность была навсегда, пока не кончилась? Разумеется, живые организмы в ходе эволюции выработали стратегии, позволяющие реагировать на непредсказуемость окружающих условий в будущем.
Самая простая и очевидная из этих стратегий — пластичность, то есть способность подстраивать свое развитие под окружающие обстоятельства. Если растение периодически теребить рукой, имитируя ветер или наличие преграды, оно вырастет невысоким и распластанным по земле, если же оставить его в покое, то оно вытянется ввысь. Разделенные близнецы могут вырасти очень непохожими друг на друга. И так далее. Итак, организмы пластичны , и это прекрасно, но есть, однако, и некоторые проблемы.
Во-первых, пластичность может быть затратной сама по себе. Все-таки, чтобы один и тот же организм мог идти по самым разным путям развития, в его геноме, клетках, органах и т. д. уже должна существовать машинерия для реализации этих направлений развития. Хорошо, если ребенок может стать и скрипачом, и физиком, но для этого он должен довольно рано изучить хотя бы азы того и другого.
Вторая проблема заключается в том, что бывают такие условия (которые могут длиться целыми эпохами), когда на свет лучше не появляться вовсе: никакая пластичность не поможет. Пропустить пару поколений и явиться в мир уже после XX съезда: многие отдали бы полжизни за такую возможность. Конечно, для людей это звучит фантастично, а вот для организмов с покоящимися стадиями (спорами, цистами, семенами) — вполне нормально. Скажем, в засушливых районах нашей планеты бывают сезоны, когда осадков так мало, что у проросших семян очень мало шансов развиться в полноценное растение. В такой ситуации возможность пропустить сезон-другой, не прорастая, может спасти гены растения. Стратегия, позволяющая избежать необратимых потерь при стрессовых или необычных условиях, пусть за счет чуть меньшего успеха в условиях «типичных», называется бет-хеджингом (см. Bet hedging). Я расскажу о ней подробнее чуть ниже, а пока…
Немного этимологии
Бет-хеджинг — заимствованный из английского языка термин, составленный из двух слов. Слово «bet» означает «ставка», в том самом смысле, в котором его употребляют букмекеры или крупье, а слово «hedging» происходит от «hedge» — «изгородь, ограда» и означает, в том числе, и ограждение себя от рисков или опасности. «Бет-хеджинг» — реально существующее понятие в лексиконе людей, увлеченных ставками на спорт. Это попытка снизить потери в случае непредвиденного результата, но сделать это малой кровью, не потратив слишком много денег. Например, некий бедолага ставит на победу фаворита в хоккейном матче довольно большую сумму. Начинается третий период, и команда, на которую ставит наш герой, ведет в счете 3:2. Вроде бы, всё идет хорошо, но в хоккее многое зависит от случайности, и он решает подстраховаться: ставит небольшую сумму на ничью, то есть фактически играет против себя, но с хорошим коэффициентом. Ведь фаворит ведет, матч почти закончен, и букмекеры дают за ничью неплохие деньги. Если матч заканчивается победой фаворита, как и ожидалось, то игрок получает выигрыш за вычетом ставки на ничью. Если же матч заканчивается вничью, то он компенсирует потери от первой ставки за счет второй и тоже теоретически — подчеркиваю: теоретически — не остается внакладе.
Возможно, слово «хедж» уже встречалось кому-то из читателей и в другом контексте. Хедж-фонды — инвестиционные организации, использующие сложные торговые стратегии для того, чтобы минимизировать риски, сохраняя при этом доходность. Вообще, хеджирование — это попытка уменьшить опасность финансовых потерь при игре на бирже, часто за счет одновременной игры и на повышение, и на понижение, чтобы подстраховаться на случай непредсказуемого поведения рынков.
По совести говоря, мне не нравится термин «бет-хеджинг» и его перевод «хеджирование ставок», абсолютно ничего не говорящие русскому уху. Однако общепринятого и подходящего перевода не существует, да и используется в русскоязычной литературе этот термин прискорбно редко. Прискорбно, потому что в последнее время бет-хеджинг в моде, и свежие статьи на эту тему довольно регулярно появляются в ведущих эволюционистских и экологических журналах (см. В изменчивом мире для успешности альтруизма достаточно, чтобы помощь была полезной в «плохие» годы, «Элементы», 12.03.2018 и Случайная изменчивость генной экспрессии у Arabidopsis thaliana подчиняется строгим закономерностям, «Элементы», 29.01.2019). Сам я предпочитаю называть бет-хеджинг «распылением ставок», чтобы подчеркнуть, что при использовании этой стратегии организм делает ставки на разные, зачастую несовместимые события в будущем (скажем, лето не может быть одновременно засушливым и дождливым), но не уверен, что мой вольный перевод придется всем по вкусу.
Бет-хеджинг в биологическом смысле: «лучше меньше, да лучше»
Говоря строго, бет-хеджинг — это стратегия, позволяющая снизить дисперсию (вариацию) собственного репродуктивного успеха за счет уменьшения среднего репродуктивного успеха. Причем если говорить совсем уж формально, то речь идет об уменьшении геометрического среднего. Я думаю, нет смысла вдаваться во все эти детали, поэтому переформулирую определение чуть более вольно. Бет-хеджинг — это стремление организма уменьшить возможный разброс числа успешных потомков за счет снижения средней численности этих потомков.
Представьте себе два растения: одно из них за три репродуктивных сезона произвело 4 + 3 + 4 взошедших семени, а второе — 100 + 0 + 0. Заметно, что одно из этих растений дает стабильный, но небольшой приплод, а у другого то густо, то пусто. Кажется, что второе растение должно оказаться в эволюционном выигрыше, поскольку в целом производит больше потомков, но давайте представим себе, что в первый год условия были благоприятные, а во второй и третий — засушливые. Тогда может так получиться, что потомки второго растения (вспомним, что они унаследовали гены своего родителя!) дадут мало потомства или не дадут никакого, в то время как потомки первого растения будут потихоньку размножаться. Тогда картина становится чуть менее однозначной. Если же предположить, что наши растения короткоживущи, а засушливые годы продолжатся, то второе растение и все его потомки могут вообще потерпеть эволюционную катастрофу, и их линия вымрет. Таким образом, может оказаться, что менее плодовитое, но более приспособленное к различным окружающим условиям растение может оказаться фаворитом на долгой эволюционной дистанции.
Каким же образом первое растение может добиться такой впечатляющей стабильности? Существуют как минимум две стратегии бет-хеджинга (см. Olofsson et al., 2009. Bet-hedging as an evolutionary game: the trade-off between egg size and number):
1) Первая — разносторонний бет-хеджинг (Diversified bet hedging), направленный на диверсификацию потомства, то есть на то, чтобы сделать его максимально разнообразным. Скажем, растение может производить два типа семян. Одни из них хорошо выживают в засушливые годы, а другие в дождливые. В результате хотя бы часть семян всходит всегда, вне зависимости от того, каким выдался год. Другой способ решить вопрос «хороших» и «плохих» годов — производить семена с разной толщиной кожуры. Семена, у которых покровы тонкие, взойдут почти все, причем уже в этом году, но если условия выдадутся неблагоприятные, то все и погибнут. Семена с толстой кожурой могут пролежать в земле несколько лет, прежде чем прорасти. Конечно, не все из них взойдут в силу разных причин: зернам кажется песок камнем, писал поэт, однако существует довольно высокая вероятность того, что хотя бы часть из них прорастет в благоприятных условиях. Многие растения действительно используют подобную стратегию (см. рис. 2). Итак, этот тип бет-хеджинга можно назвать в соответствии с англо-саксонской поговоркой «Не надо складывать все яйца в одну корзину». Ну или, в соответствии с китайским изречением, «Пусть растут все цветы».
Наиболее совершенная форма диверсифицированного бет-хеджинга называется «адаптивное подбрасывание монетки» (Adaptive coin flipping). Представьте себе многолетнее растение, растущее в засушливом районе и год от года производящее семена. Вместо того чтобы просто давать семена разных типов — авось кто-то выживет, — растение поступает хитрее. Каждый год оно «бросает монетку» и случайным образом выбирает, к какому типу будет относиться каждое из его семян: тому, что хорошо приспособлен к засушливым годам, или тому, что хорошо прорастает в дождливую погоду. Важно, что вероятность выбора каждой стратегии соответствует вероятности того, что год будет дождливым или засушливым. Если вероятность дождливого года 25%, то примерно четверть семян и будут приспособлены к таким условиям. Конечно, в некоторых случаях выбор будет не совсем удачный, и растение проиграет «чистым» специалистам по той или иной стратегии, но в долгосрочной перспективе подбрасывание монетки работает лучше любой из них.
2) Вторая стратегия — консервативный бет-хеджинг (Conservative bet hedging). Ее можно назвать «Плохонький, да мой», или же «Ни рыба ни мясо», или «Тише едешь, дальше будешь»). Она заключается в том, чтобы, напротив, оставить однотипное потомство, имеющее, однако, хорошие шансы выжить в любых условиях. То есть это будет нечто среднее, неспециализированное, универсальное, без особого эволюционного полета, но очень неприхотливое. В этом случае организм не пытается диверсифицировать стратегии выживания своих потомков, а, напротив, делает ставку на одну и ту же безопасную стратегию.

Рис. 2. Схема, иллюстрирующая успешность бет-хеджинга по сравнению со специализированными стратегиями. Хорошо заметно, что специалисты оказываются в выигрыше, когда условия окружающей среды стабильны: подавляющая часть годов либо хорошая, либо плохая. Если же условия непредсказуемы, то стратегия бет-хеджинга может оказаться наиболее успешной. Особенно важно то, что бет-хеджинг в любом случае дает более предсказуемые результаты, то есть всегда работает приемлемо (обратите внимание, что кривые зависимости приспособленности от доли «хороших» годов почти параллельны оси Х). На врезках — примеры бет-хеджинга: черно-белая врезка — два типа семян однолетнего растения Heterotheca latifolia, использующие разные стратегии распространения и прорастания (верхний ряд — общий вид, нижний — поперечный срез). Те, что слева, хорошо переносятся ветром и быстро прорастают, а те, что справа, — не летучие, с утолщенными покровами для пережидания неблагоприятных условий. Цветная врезка: покоящиеся яйца дафний, способные столетиями лежать на дне, дожидаясь подходящего момента для вылупления рачков. Источники изображений: en.wikipedia.org, M. E. Evans, J. J. Dennehy, 2005. Germ banking: bet-hedging and variable release from egg and seed dormancy и ezramagazine.cornell.edu, с изменениями
Долгое время основным объектом исследования бет-хеджинга были однолетние растения, живущие в пустынях и полупустынях. Со временем выяснилось, что и другие живые существа могут использовать бет-хеджинг при формировании своей стратегии размножения (см. M. E. Evans, J. J. Dennehy, 2005. Germ banking: bet-hedging and variable release from egg and seed dormancy). Так, размер выводка у птиц может определяться желанием родителей подстраховаться на случай неподходящих условий. Покоящиеся яйца многих планктонных животных, способные сотни лет пролежать в иле, тоже могут считаться попыткой распылить ставки. В последнее время выяснилось, что и вирусы не пренебрегают бет-хеджингом.
В своей работе группа американских ученых выяснила, какие механизмы ответственны за формирование стратегии бет-хеджинга у цитомегаловируса (ЦМВ) — одного из самых распространенных вирусов в человеческой популяции (рис. 1).
Вирионы цитомегаловируса: оппортунисты и радикалы
Цитомегаловирус, также известный под названием вирус герпеса человека 5-го типа, представляет собой двухцепочечную ДНК, погруженную в плотную белковую оболочку — капсид — и окруженную тегументом. ДНК этого вируса содержит довольно много генетического материала, в котором закодировано много вирусных белков, а потому цитомегаловирус имеет весьма гибкий жизненный цикл (рис. 3), в ходе которого может переключаться с одной стратегии эксплуатации хозяина на другую .


Рис. 3. Упрощенная схема литического цикла цитомегаловируса в клетке. Частицы вируса проникают в клетку, оставляя часть белков своего тегумента, включая pp71, в цитоплазме. Затем ДНК вируса поступает в ядро, где с него считывается множество копий. Уже в ядре формируется капсид вируса, вокруг которого в цитоплазме клетки-хозяина собираются белки тегумента и гликопротеины мембраны. После этого новорожденные вирионы покидают клетку, готовые к новым свершениям. На врезке: схема устройства вирусной частицы ЦМВ. Источники изображений: T. Crough and R. Hanna, 2009. Immunobiology of Human Cytomegalovirus: from Bench to Bedside и T. Manandhar et al., 2019. Battle between Host Immune Cellular Responses and HCMV Immune Evasion, с изменениями
ЦМВ способен переноситься самыми разными путями, включая передачу через слюну, при переливании крови, половом контакте, а также переходить от матери к ребенку еще в утробе или при кормлении грудью (T. Crough and R. Hanna, 2009. Immunobiology of Human Cytomegalovirus: from Bench to Bedside). Поэтому неудивительно, что этим вирусом заражена большая часть человечества. ЦМВ вообще-то вирус мягкий, предпочитающий не вступать в жесткую конфронтацию со своим хозяином. Инфекция, как правило, протекает бессимптомно, реже ее первичное проявление ограничивается мышечной болью, тонзиллитом, небольшим жаром и/или легким воспалением печени. После этого вирус остается в организме человека, но надолго уходит в тень, лишь периодически напоминая о себе, пользуясь временным ослаблением иммунитета хозяина из-за других инфекций, стресса или воспаления (T. Crough and R. Hanna, 2009). Однако существуют категории граждан, для которых ЦМВ может представлять серьезную угрозу. Обычно это люди с ослабленным или еще не сформировавшимся иммунитетом, например недавно перенесшие трансплантацию органов, носители ВИЧ и новорожденные. Считается, что ЦМВ — основная причина врожденных неврологических расстройств (таких, например, как глухота), а порой цитомегаловирусная инфекция может привести и к смерти ребенка. Хотя случаи тяжелого течения инфекции довольно редки, широкое распространение вируса в человеческой популяции приводит к заметному числу жертв ЦМВ в абсолютном выражении.
Вирус проникает в клетку человека, взаимодействуя с рецепторами клетки хозяина с помощью гликопротеинов на поверхности своего тегумента. После проникновения белки его тегумента и капсида используют микротрубочки хозяина, чтобы доставить ДНК вируса в ядро клетки. В ходе этого продвижения вирус начинает «раздеваться», постепенно теряя белки своей оболочки в цитоплазме. «Потерянные» белки не тратят время понапрасну, взаимодействуя с различными структурами в клетке, чтобы помешать развитию иммунного ответа хозяина. Затем ДНК вируса проникает в ядро и коварно использует синтетический аппарат клетки хозяина для создания собственных копий и белков, необходимых для успешного формирования вирусных частиц. Сформировавшиеся de novo капсиды вируса поступают из ядра в цитоплазму, где, опять же с использованием клеточных структур хозяина, одеваются тегументом, после чего выходят наружу заражать другие клетки. Внешний вид зараженных клеток заметно меняется: они увеличиваются в размерах (отсюда название вируса), а в ядре появляются характерные включения, хорошо заметные при окраске гематоксилином и эозином. Я, разумеется, опустил массу интересных деталей и подробностей, но даже в таком куцем виде жизненный цикл вируса не лишен определенного изящества. То, что я сейчас описал, это лишь активная форма (литический путь) существования вируса, в которой вирус быстро воспроизводится.
Отличительной особенностью герпесвирусов (группы, к которой ЦМВ относится) является способность переходить в латентное, «спящее», состояние (S. H. Speck and D. Ganem, 2010. Viral latency and its regulation: lessons from the gammaherpesviruses). При необходимости, а необходимость возникает довольно часто , ЦМВ способен задраить все люки, лечь на дно и сделаться невидимым для сонаров иммунной системы хозяина, терпеливо выжидая момента, когда ему представится шанс вновь приступить к активному размножению. В таком случае ДНК цитомегаловируса «молчит», просто находясь в ядре клетки хозяина , и вирус ждет своего часа. Вы, я думаю, не понаслышке знаете, что и обычный герпес ведет себя сходным образом. В регуляции того, будет вирус активен или перейдет в спящее состояние, важную играют роль два белка: pp71 и pp150, причем первый из них активирует белки, отвечающие за копирование вирусного генома, а второй, напротив, подавляет их работу, переводя ЦМВ в латентное состояние. Конечно, есть и другие регуляторные белки, но авторы исследования в первую очередь акцентировали внимание именно на этих двух.
Каким же образом генетически идентичные вирусные частицы могут различаться по стратегиям взаимодействия с клеткой хозяина? Как определяется, какая частица будет активной, а какая латентной? Вот здесь и выступает на сцену бет-хеджинг.
Бет-хеджинг на вирусный манер
Чтобы придумать, как обнаружить бет-хеджинг у ЦМВ, и показать, как он работает, группе американских исследователей пришлось решить последовательно несколько задач. Во-первых, им нужно было узнать, есть ли существенные различия в количестве молекул белков рp71 и pp150, отвечающих за переход в спящий режим , в разных частицах вируса (по аналогии с семенами растений: если у какого-нибудь вида вы обнаруживаете сильную вариацию в толщине покровов, то это может навести вас на мысль, что растение использует бет-хеджинг, чтобы приноровиться к разным условиям ). Для этого использовалась флуоресцентная микроскопия сверхвысокого разрешения (см. Super-resolution microscopy): вирусы, как правило, настолько малы, что в обычный световой микроскоп их не увидишь. Я не буду здесь рассказывать о том, что такое микроскопия сверхвысокого разрешения, — это увело бы нас слишком далеко от темы сегодняшнего разговора, — зато в общих чертах изложу сущность флуоресцентного метода.
Исследователи пометили интересующие их белки ЦМВ флуоресцентной меткой, «пришив» к ним желтый флуоресцирующий белок (YFP — yellow fluorescent protein). Из-за этого под воздействием света определенной длины волны молекулы pp71 и pp150 начинали флуоресцировать, испуская cвет с длиной волны около 527 нм. Поскольку YFP излучает в относительно узком диапазоне длин волн, весь «лишний» свет можно убрать с помощью специальных фильтров, оставив только специфическое излучение, исходящее от интересующих исследователей белков, а интенсивность этого излучения измерить. Хорошая новость заключается в том, что с увеличением числа молекул меченого белка внутри вириона (вирусной частицы) почти линейно увеличивается и интенсивность свечения. А плохая в том, что чувствительность флуориметров, свойства фильтров и программное обеспечение по обработке изображений заметно разнятся, затрудняя измерение интенсивности флуоресценции в абсолютных величинах. Конечно, эту проблему можно решить калибровкой и стандартизацией приборов, но биологи в своих исследованиях часто «не парятся» этим, поскольку для них бывает важнее понять, насколько разные группы клеток (скажем, клетки в опыте и контроле) отличаются по содержанию белка друг от друга, то есть их интересует относительная, а не абсолютная интенсивность свечения. В таком случае результат измерения интенсивности считают безразмерной величиной и сообщают в произвольных или условных единицах (a.u., arbitrary units), заставляющих тех из нас, кто постарше, вспомнить лихие 90-е и времена галопирующей инфляции. Поэтому пусть читателей не смущает, что на рисунках в статье интенсивность представлена как безразмерная величина. Убрав весь ненужный свет с помощью фильтров, исследователи получали картинки (см. рис. 4), на которых частицы ЦМВ были видны как зеленые пятна на темном фоне. Эти изображения затем анализировались.

Рис. 4. A. Слева: схема внутреннего устройства вириона цитомегаловируса. Показаны ассоциированный с капсидом белок pp150 во внутреннем тегументе и белок pp71 во внешнем слое тегумента. Справа: вирионы ЦМВ, содержащие белки pp71 и pp150, помеченные желтым флуоресцентным белком (ЖФБ = YFP). Разброс в интенсивности свечения заметен даже невооруженным взглядом. И таки да, это нормально, что он зеленый , несмотря на название. B, C. Распределение частиц вируса по интенсивности свечения ЖФБ, то есть по содержанию меченых белков pp71 и pp150. Очевидно, что разброс интенсивностей в случае с pp71 (зеленый) гораздо больше, чем у белка pp150 (синий) и одного из белков капсида вируса «обычного» герпеса, взятого в качестве своего рода контроля (черный). Здесь и далее рисунки из обсуждаемой статьи Chaturvedi et al., 2020, с изменениями
В ходе анализа выяснилось, что число молекул белка pp71 в разных клетках действительно, сильно отличалось, причем разброс был гораздо больше, чем у белка pp150 или белка, взятого в качестве контроля. Более того, оказалось, что различия между клетками по интенсивности средней флуоресценции вирусных частиц невелики, а значит, за разброс были ответственны именно различия между частицами вируса. Другими словами, исследователи исключили вероятность того, что на концентрацию белка pp71 могут заметно повлиять клетки-хозяева. Всё говорило о том, что вирус целенаправленно способствует синтезу частиц с разным количеством этого белка. Напомню, что белок pp71, как предполагалось, отвечает за активацию более агрессивной стратегии цитомегаловируса, заключающейся в активном производстве частиц вируса в ядре и цитоплазме клеток хозяина.
На следующем этапе необходимо было выяснить, какие именно эволюционные преимущества и/или недостатки кроются за различными концентрациями pp71. Ученые использовали линию клеток-хозяев, в которых ген вируса, кодирующий белок pp71, экспрессировался сильнее для создания вирусных частиц с повышенным содержанием этого белка, и затем сравнили их репродуктивный успех с успехом вирусных частиц, содержащих «обычное» количество pp71 (обозначены на графиках ниже как pp71HI и pp71WT соответственно). В качестве меры репродуктивного успеха авторы статьи использовали такой показатель, как инфективность вируса (см. Infectivity), то есть способность его частиц успешно заражать клетки хозяина и размножаться в них. Для этого определяли функциональный титр вируса и измеряли количество копий вируса ДНК в клетках. Не вдаваясь в подробности, скажу, что функциональный титр можно интерпретировать как число вирусных частиц, способных успешно размножиться в клетке хозяина. Для того чтобы его рассчитать, определяют объем содержащего вирус раствора, необходимый для того, чтобы заразить определенное количество хозяев — скажем, половину мышей, крыс или добровольцев-людей, подвергнувшихся воздействию инфекции.
В случае с ЦМВ в качестве «подопытных» выступали культуры клеток. Зная титр вируса, можно рассчитать и примерное число вирусных частиц в объеме раствора, способных успешно размножиться в клетке хозяина. При этом истинное число вирусных частиц в растворе может быть гораздо больше, то есть, в каком-то смысле, инфекционный титр измеряет не столько число вирусных частиц, сколько «агрессивность» вируса. Часто титр вируса выражают в виде числа бляшкообразующих единиц в единице объема (PFU, plaque forming units / ml на графике внизу). Давайте для краткости заметем под ковер, откуда на графике взялись бляшкообразующие единицы и что это такое. Просто примем, что каждая из них соответствует одному или группе вирионов, успешно проникших в клетку и пошедших по литическому пути развития. Итак, чем выше концентрация таких вирусных частиц, тем выше и титр, то есть чем больше титр вируса, тем более вирус инфективен. Другими словами, если мы возьмем два раствора, содержащих одинаковое количество частиц вирусов А и Б, и добавим их к клеточной культуре, то более инфективный вирус сможет заразить больше клеток за заданное время. И наоборот если мы возьмем одинаковые по инфективности растворы вирусов, то ДНК более «успешного» вируса будет содержаться в этом растворе в меньшем количестве: раз вирус более эффективно заражает клетки, то и нужно его меньше.
Оказалось, что вирусные частицы с повышенным содержанием pp71 (pp71HI) активно размножаются в клетках хозяина в разы чаще, чем вирионы с нормальным количеством этого белка (pp71WT). Вирусные частицы, богатые pp71, заражали больше клеток в культуре, чем то же количество нормальных частиц, и их требовалось меньше для достижения сходной эффективности заражения (рис. 5).

Рис. 5. A. Интенсивность флуоресценции (= количество) белка pp71 в частицах вируса, полученных в обычных клетках хозяина и клетках, где ген вируса, отвечающий за синтез этого белка, экспрессировался сильнее. Красный квадрат — средняя интенсивность свечения вирусных частиц. Хорошо заметно, что интенсивность свечения вирионов pp71HI заметно выше, чем у вирионов «дикого типа» (WT) — в первую очередь, за счет относительно небольшого числа вирусных частиц с особенно большим содержанием pp71. B. Справа: при одинаковом числе частиц вируса в растворе инфективность вирионов с повышенным количеством белка pp71 была выше. Слева: и наоборот, когда растворы выравнивали по инфективности, число копий более «агрессивных» вирионов с большим количеством белка pp71 было ниже. C. Интенсивность флуоресценции белка IE2 (см. ниже), помеченного ЖФБ, свидетельствует о том, что вирус в клетке пошел по литическому пути развития. Хорошо заметно, что в обычных фибробластах (клетках соединительной ткани) такой белок начинал синтезироваться реже, чем в фибробластах, где ген вируса, отвечающий за синтез белка pp71, экспрессировался сильнее. Неудивительно, что «выращивание» вируса на клетках этой линии в течении семи дней приводило к получению более инфективных частиц (D)
Однако и на Солнце есть пятна…
…которые исследователи попытались обнаружить на следующем этапе. Рассуждали они так: если вирионы с большим количество белка pp71 так хороши, то почему не все вирусные частицы содержат его так много? Казалось бы, естественный отбор должен был всячески этому благоприятствовать. Почему же линии вируса с высоким содержанием этого активирующего размножение вируса белка не вытеснили своих менее агрессивных конкурентов? Дело в том, что вирусное счастье заключается не только в заражении максимального числа клеток. Подобно преступникам, постоянно и демонстративно нарушающим закон, слишком активные вирусы чересчур быстро привлекают внимание силовых ведомств организма и в результате не живут долго. Слишком активная подрывная деятельность вируса внутри тела хозяина чревата активным иммунным ответом, а, как уже отмечалось выше, в здоровом организме ЦМВ предпочитает проводить большую часть времени «в тени», лишь изредка переходя границы дозволенного.
Как выяснилось, именно избыток белков pp71 мешает вирусным частицам перейти на легальное положение. Механизм происходящего, видимо, таков. Проникая внутрь ядра клетки хозяина, ДНК цитомегаловируса деликатно оставляет за дверью, то есть в цитоплазме, большую часть белков своего капсида и тегумента, в том числе и белок pp71. Тем не менее некоторое его количество все-таки проникает в ядро, и то, по какому пути пойдет развитие инфекции — литическому (пути активного размножения вируса) или латентному, — зависит именно от концентрации белка pp71 в ядре клетки. Если его много, то вероятность перехода вируса в спящее состояние невелика.
Для того чтобы показать это экспериментально, авторы статьи проверяли, как присутствие повышенных количеств pp71 в ядре влияет на экспрессию генов, отвечающих за развитие литического пути размножения вируса. Если вирус выбрал именно этот способ действия в организме хозяина, то вскоре после проникновения в клетку-хозяина в его геноме начинают работать гены раннего ответа (Immediate Early genes). Считываемые с них белки так и называются IE1 и IE2. Если этих белков в клетке мало, то есть весомые основания предполагать, что вирус предпочел не проявлять заметной внутриклеточной активности и перейти в спящий режим. Исследователи «подсветили» белки IE1 и IE2 двумя разными видами флуоресцентных меток и проверили, насколько сильно экспрессируются гены раннего ответа в клетках, зараженных вирионами, содержащими разное количество белка pp71. Оказалось, что лишь клетки, в которых концентрация этого белка очень велика, продуцировали большое число белков раннего ответа, а в остальных вирус, видимо, переходил в латентное состояние.
Наконец, авторы статьи напрямую проверили гипотезу о том, что вирусные частицы с высоким содержанием белка pp71 хуже способны «засыпать» в клетках хозяина (рис. 6). Исследователи заражали клетки вирусом с нормальным и высоким содержанием белка рр71, ждали четыре дня, пока вирус перейдет в латентное состояние, а потом реактивировали его. Выяснилось, что в клетках, зараженных вирусом с повышенным содержанием pp71, вирус реже оказывался способен задерживаться надолго: меньшее число таких клеток содержало вирус спустя несколько дней после заражения, и в меньшей их части он успешно возвращался к литическому пути развития при реактивации. Другими словами, в долгосрочной перспективе вирусные частицы, богатые белком pp71, оказывались хуже приспособлены к переходу в спящий режим с последующей реактивацией, чем их менее «честолюбивые» (при первой встрече с хозяином) собратья.

Рис. 6. Вверху справа: количество белка pp71 в ядре зараженных клеток сильно коррелировало с интенсивностью экспрессии вирусных белков раннего ответа, что свидетельствует о том, что избыток pp71 может препятствовать переходу вируса в спящий режим. Обратите внимание (слева вверху), что в клетках зараженных «диким» (WT) типом ЦМВ интенсивность свечения белков раннего ответа вируса, подсвеченных желтым и красным флуоресцирующими белками, фактически не отличалась от фонового свечения, полученного в контроле, где вируса не было вообще (см. правый верхний квадрант графиков). Другими словами, почти во всех клетках, зараженных вирионами с нормальным содержанием pp71, белки раннего ответа не экспрессировались вовсе, то есть вирус перешел в латентное состояние. При этом в заметной доле клеток (~6%), зараженных вирусными частицами с избытком pp71, белки раннего ответа экспрессировались (справа внизу). Нижний ряд справа налево: меньшее число клеток разных линий (NTera2, CD14+), зараженных вирионами pp71HI, содержало вирус спустя несколько дней после заражения (30–40% от числа клеток, зараженных вирусом с обычным количеством этого белка (рр71WT), и в меньшей их части вирус успешно возвращался к литическому пути развития при реактивации
Что в итоге?
Таким образом, оказалось, что как большое, так и малое число молекул белка pp71 в тегументе цитомегаловируса имеет свои достоинства и недостатки в зависимости от обстоятельств. Именно поэтому вирусу может оказаться выгодно помещать разные количества этого белка в генетически идентичные вирионы. Это позволяет ЦМВ быть более гибким в экологическом смысле и использовать разные стратегии эксплуатации хозяина, приноравливаясь к различным условиям окружающей среды: либо делая ставку на немедленное и активное размножение, либо выбирая более умеренную выжидательно-оппортунистическую политику. Подобно растению, производящему разные типы семян, чтобы быть готовым к различным условиям и выжить в засушливые годы даже ценой потери части потомства, вирус готов пожертвовать некоторыми из своих потомков, чтобы другие успешно размножились и продолжили «радовать» нас тонзиллитом, жаром, болью в мышцах, а иногда и куда более заметными и неприятными симптомами. Таков бет-хеджинг в вирусном исполнении!
Авторы статьи провели очень интересное и детальное исследование различных репродуктивных стратегий цитомегаловируса, продемонстрировали любопытный пример бет-хеджинга у вирусов и показали, что базовые эволюционно-экологические стратегии могут сходным образом работать как в макро- так и в микромире.
Источник: elementy.ru





