













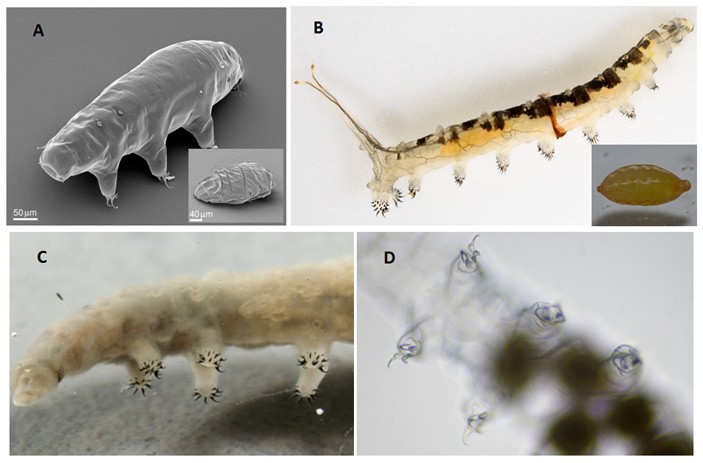

Рис. 1. Морфологическое сходство тихоходок и личинок экстремофильных мух семейства Ephydridae. А — тихоходка Milnesium tardigradum (на врезке — дегидратированная особь в состоянии криптобиоза), B — личинка солелюбивой мухи Cirrula hians (= Ephydra hians) из соленого щелочного озера Моно в Калифорнии (на врезке — куколка), C — псевдоподии личинки Ephydra cinerea из Большого Соленого озера, D — лобоподии тихоходки Tardigrada gen. et sp. indet. Изображение из дополнительных материалов к обсуждаемой статье, фото с сайтов researchgate.net, bugguide.net, esa.int
Происхождение тихоходок — мелких животных, известных своей устойчивостью к экстремальным условиям среды, — долго оставалось загадочным, хотя их родство с членистоногими признавалось всеми специалистами. Новые методы молекулярно-филогенетического анализа наряду с морфологическими данными показали, что тихоходки, по-видимому, являются неотеническими личинками двукрылых насекомых семейства Ephydridae (мушки-береговушки). К признакам, общим для тихоходок и личинок эфидрид, относятся исключительная выносливость (эфидриды способны развиваться в пересоленной воде, при экстремальных значениях pH, в сырой нефти и даже в скипидаре), рост с линьками, сходное строение примитивных конечностей — лобоподий, а также коготков на них. Обработка тихоходок гормонами, стимулирующими окукливание эфидрид, приводит к переходу животных при последующем высушивании в состояние криптобиоза, которое, по-видимому, гомологично стадии куколки обычных двукрылых. Многие уникальные особенности тихоходок, такие как сокращение числа сегментов тела и утрата дыхательной (трахейной) системы, по-видимому, являются следствием миниатюризации. Полученные результаты объясняют, почему тихоходки лишились трех Hox-генов (Scr, Antp и Ubx), которые у других насекомых отвечают за формирование грудных сегментов. У личинок эфидрид грудные сегменты, в отличие от брюшных, никогда не несут конечностей, поэтому при неотении и миниатюризации они оказались бесполезными и были утрачены.
Тихоходки (Tardigrada) — группа, интересная во многих отношениях. Широкой публике они известны в основном из-за своего неотразимого очарования. Недаром их называют «медвежатками», «водяными мишками» (water bears) и «моховыми поросятами» (moss piglets). Кроме того, тихоходки славятся своей невероятной выносливостью. Например, экспериментально показано, что они выдерживают 10 суток пребывания в открытом космосе (K. I. Jönsson et al., 2008. Tardigrades survive exposure to space in low Earth orbit). Неудивительно, что тихоходки стали героями сетевых мемов (рис. 2).

Рис. 2. Сетевой мем, прославляющий выносливость тихоходок. Изображение с сайта beatricebiologist.com
Живучесть тихоходок во многом определяется их способностью переносить почти полное обезвоживание (дегидратацию), впадая в состояние криптобиоза (рис. 1, А, на врезке). В этом состоянии тихоходкам не страшны ни космический вакуум, ни большие дозы радиации, ни замораживание в жидком гелии, ни кипячение, ни давление в 6000 атмосфер.
Происхождение тихоходок — давняя научная загадка. По многим признакам тихоходки явно близки к членистоногим, онихофорам, а также к некоторым ископаемым базальным представителям надтипа Ecdysozoa (см. Линяющие), известным по находкам из кембрийских лагерштеттов. Однако точное положение тихоходок в системе Ecdysozoa долго не удавалось установить.
Новое исследование американских биологов, изучающих галофильную фауну Большого Соленого озера в штате Юта (США), проливает неожиданный свет на эту проблему.
Ученые обратили внимание на морфологическое сходство конечностей тихоходок и личинок солелюбивых мух-береговушек (семейство Ephydridae), обитающих в Большом Соленом озере (рис. 1, C, D). Для тихоходок характерно наличие четырех пар мягких нечленистых конечностей — лобоподий (см. lobopod), несущих характерные изогнутые коготки. Очень похожие конечности и коготки были у некоторых кембрийских родственников членистоногих, таких как галлюцигения (см.: Коготки галлюцигении подтвердили гипотезу о происхождении членистоногих, «Элементы», 21.10.2014).
У большинства двукрылых насекомых (комаров и мух) личинки не имеют конечностей. Однако у обитающих в Большом Соленом озере личинок мушек-береговушек Ephydra cinerea, как и у многих других представителей семейства Ephydridae, на брюшных сегментах имеется семь пар псевдоподий (ложноножек), внешне практически неотличимых от конечностей тихоходок. Три грудных сегмента, на которых у взрослой мухи находятся настоящие членистые ноги, у личинок не несут конечностей (как и положено личинкам мух).
Как объяснить это сходство? На первый взгляд, проще всего предположить конвергентную эволюцию, то есть независимое формирование похожих конечностей у организмов, живущих в сходных условиях. И тихоходки, и личинки эфидрид живут в водной среде, используя свои мягкие когтистые ножки для ползания по различным субстратам. Но такое простое объяснение не удовлетворило исследователей, решивших, что вопрос заслуживает более серьезного изучения.
Ученые применили к имеющимся в генбанке нуклеотидным последовательностям тихоходок и эфидрид новый оригинальный метод построения филогенетических деревьев, использовав в качестве внешней группы (см. Outgroup) родственное эфидридам семейство Drosophilidae. У дрозофил, кстати, личинки безногие, как и у большинства других мух. Получившееся дерево подтвердило гипотезу о родстве эфидрид и тихоходок (рис. 3).

Рис. 3. Эволюционное дерево тихоходок, показывающее их родство с мушками-береговушками (Ephydridae). При построении дерева в качестве внешней группы использовалось родственное эфидридам семейство дрозофилид. Буквами обозначены места обитания тихоходок: M — морские семейства, T — наземные, F — пресноводные. Изображение из обсуждаемой статьи (по данным из статьи P. B. Nichols et al., 2006. A Family Level Analysis of Tardigrade Phylogeny, с изменениями)
Помимо морфологического сходства и результатов молекулярно-филогенетического анализа, в пользу этой гипотезы свидетельствуют и другие факты. Авторы скрупулезно разбирают их в своей статье, уделяя особое внимание двум обстоятельствам.
Во-первых, ранее было показано, что характерная для тихоходок способность выдерживать обезвоживание и пережидать тяжелые времена в состоянии криптобиоза встречается также и у личинок двукрылых (M. Watanabe et al., 2003. Increase of internal ion concentration triggers trehalose synthesis associated with cryptobiosis in larvae of Polypedilum vanderplanki). По-видимому, эта способность унаследована ими от общего с тихоходками предка.
Во-вторых, личинки эфидрид, как и тихоходки, отличаются невероятной выносливостью. Например, личинки мухи Cirrula hians (другое название — Ephydra hians, рис. 1, B) чувствуют себя вполне комфортно в пересоленной щелочной воде озера Моно в Калифорнии, в которой не может выжить ни одна рыба (см. рисунок 2 в новости Симбиотические дрожжи помогают дрозофилам адаптироваться к соленому корму, «Элементы», 11.11.2019). Личинки другого вида эфидрид, Helaeomyia petrolei, развиваются в естественных лужах сырой нефти. Кишечник у них всегда полон нефти, хотя питаются они на самом деле не нефтью, а упавшими в лужу насекомыми. Лабораторные опыты показали, что с тем же успехом эти личинки могут жить в 50-процентом скипидаре (см. картинку дня Нефтяная муха). Физиологические и биохимические механизмы устойчивости тихоходок и эфидрид к неблагоприятным внешним воздействиям еще не до конца изучены. Но с большой вероятностью она имеет единое происхождение: трудно предположить, что такая уникальная особенность могла возникнуть два раза независимо.
На основании всех этих фактов ученые заключили, что тихоходки, скорее всего, произошли от эфидрид путем неотении. Иными словами, тихоходки — это потомки мух-береговушек, перешедшие к размножению на личиночной стадии и утратившие стадию имаго (взрослого крылатого насекомого).
Неотения — довольно частое явление в эволюции (см. Что общего у голых землекопов и «голых обезьян»?, «Элементы», 06.03.2017). Хотя самые известные примеры неотении связаны с амфибиями (см. Неотения у амбистомовых), у насекомых она тоже встречается (M. Bocakova et al., 2007. Molecular phylogenetics of Elateriformia (Coleoptera): evolution of bioluminescence and neoteny). Так что неотенические личинки эфидрид в этом плане не являются исключением.
После перехода к неотении тихоходки сильно измельчали. Это позволило им отказаться от трахей и перейти к дыханию всей поверхностью тела. Именно поэтому у тихоходок отсутствует сифон — дыхательная трубка на заднем конце тела, характерная для личинок обычных (не неотенических) береговушек.
Другим следствием миниатюризации стало уменьшение числа сегментов тела (олигомеризация). С этим связано еще одно — возможно, самое красивое и убедительное — подтверждение идеи о неотенической природе тихоходок, основанное на данных сравнительной геномики и эволюционной биологии развития (evo-devo).
Ранее было показано, что олигомеризация у тихоходок была сопряжена с утратой нескольких Hox-генов (см. Укорочение тела тихоходок связано с потерей Hox-генов, «Элементы», 04.03.2016). Эти гены у билатерально-симметричных животных участвуют в продольной «разметке» эмбриона и определении судьбы сегментов. При этом оставалось неясным, почему тихоходки утратили одни Hox-гены (и, по-видимому, соответствующие им сегменты), но сохранили другие. Была ли в этом какая-то система? От каких именно сегментов или отделов тела отказались тихоходки?
Как показано на рисунке 2 в вышеупомянутой новости, три важнейших Hox-гена, утраченных тихоходками, это Sex combs reduced (Scr), Antennapedia (Antp) и Ultrabithorax (Ubx). Экспрессия этих генов у насекомых маркирует сегменты грудного отдела: Scr — переднегрудь, Antp — среднегрудь, Ubx — заднегрудь.
Пока тихоходки считались группой неясного систематического положения, а их родственные связи с насекомыми оставались спорными, дать этому факту сколько-нибудь внятную интерпретацию не удавалось. Но теперь, когда стало ясно, что тихоходки — это насекомые отряда двукрылых, всё сразу встало на свои места. Получается, что у тихоходок в процессе олигомеризации редуцировался грудной отдел тела. Самое интересное, что именно этого и следовало ожидать в случае неотении и последующей миниатюризации у мух-береговушек. Ведь у личинок мух вообще и эфидрид в частности, как мы помним, на грудных сегментах нет никаких конечностей — ни настоящих членистых ног, ни ложноножек-лобоподий (не говоря уже о крыльях). Однако брюшные сегменты у личинок эфидрид несут когтистые лобоподии, чрезвычайно удобные для ползания по самым разнообразным подводным субстратам. От каких же сегментов, грудных или брюшных, должны были в первую очередь отказаться неотенические личинки эфидрид в ходе миниатюризации? Конечно, от бесполезных грудных, что и подтверждается данными сравнительной геномики и evo-devo. Число брюшных сегментов с парными лобоподиями у тихоходок сократилось с семи до четырех, что согласуется с утратой одного из «брюшных» Hox-генов, Abd-A.
Таким образом, смелая гипотеза американских биологов не только подтвердилась многочисленными фактами, но и пролила неожиданно яркий свет на закономерности сопряженной эволюции плана строения организма и ключевых генов — регуляторов развития.
В заключительной части работы авторы обсуждают возможную гомологию стадии куколки обычных двукрылых насекомых и дегидратированного криптобиотического состояния тихоходок. Процесс впадания тихоходок в криптобиоз действительно напоминает окукливание личинок эфидрид (рис. 1, A, B), хотя есть и очевидные различия. В частности, окукливание у эфидрид не сопровождается полным обезвоживанием.
Дополнительные эксперименты показали, что если обработать тихоходок гормоном экдизоном, который стимулирует линьки и окукливание насекомых, то после высушивания тихоходки впадают в криптобиоз. Для полноты картины эти эксперименты следовало бы дополнить контрольными опытами с высушиванием, но без экдизона. Авторы пока не успели этого сделать. Данный вопрос, несомненно, заслуживает дальнейшего изучения.
Источник: elementy.ru





