













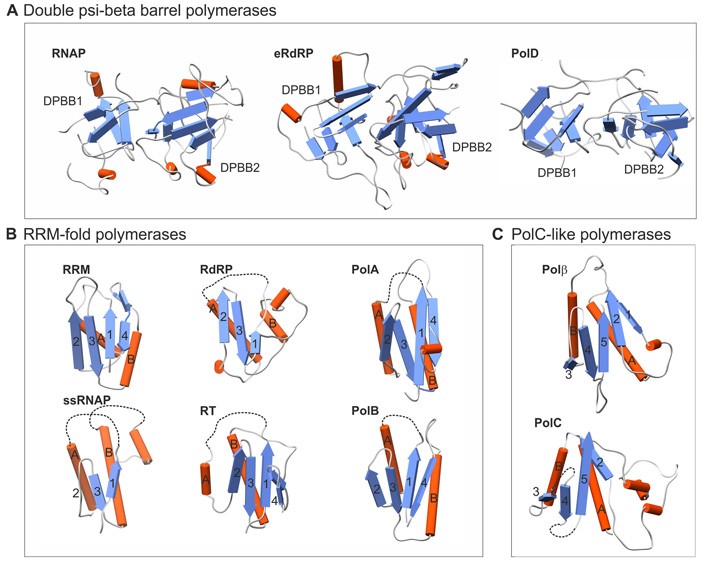


Рис. 1. Строение основных структурных мотивов, образующих активный центр ДНК- и РНК-полимераз. A — мотив двойной ψβ-бочонок (DPBB), B — мотив RRM, C — активный центр ДНК-полимераз семейства PolC. Голубыми стрелками обозначены β-листы, красными цилиндрами — α-спирали. Рисунок из обсуждаемой статьи в BMC Biology
Происхождение важнейшего молекулярно-биологического процесса, лежащего в основе размножения любой живой клетки, — репликации ДНК — окутано тайной. Дело в том, что ферменты, обеспечивающие удвоение цепочек ДНК — ДНК-полимеразы — у организмов из трех доменов жизни (архей, бактерий и эукариот) неродственны друг другу. Недавно группа исследователей во главе с Евгением Куниным предложила изящную гипотезу возникновения репликационного аппарата, согласующуюся с известными данными о сходстве и различиях ДНК-полимераз у архей, бактерий и эукариот. Ученые считают, что ДНК-полимеразы и РНК-полимеразы (ферменты, осуществляющие транскрипцию) произошли от одного предкового белка, который функционировал, как РНК-зависимая РНК-полимераза — синтезировал комплементарную РНК на матрице РНК.
Существует великое множество гипотез происхождения жизни на Земле, и многие из них сходятся на том, что в определенный период на земле существовал некий гипотетический организм, от которого произошли все ныне живущие клеточные формы жизни — последний универсальный общий предок, или LUCA (от англ. Last Universal Common Ancestor; подробнее о том, каким мог быть LUCA и почему большинство ученых уверены в его существовании, читайте в новости Формальные статистические тесты подтверждают происхождение всех живых организмов от единого предка, «Элементы», 19.05.2010).
Если все формы жизни происходят от одного общего предка, то кажется разумным, что ферменты, обеспечивающие протекание базовых биологических процессов, общих для всего живого, должны быть очень близки у всех современных организмов. В самом деле, белки, участвующие в транскрипции, трансляции и ключевых метаболических процессах вроде синтеза нуклеотидов, крайне консервативны и очень близки даже у эволюционно далеких групп организмов (в частности, — у кишечной палочки и у человека).
Однако есть одно странное исключение, которое, на первый взгляд, ставит под сомнение если не существование LUCA, то, как минимум, наличие у него генома в виде молекулы ДНК — репликация ДНК. Дело в том, что ферменты ДНК-полимеразы, которые катализируют присоединение новых дезоксирибонуклеотидов, комплементарных матричной цепи ДНК, у представителей трех доменов клеточных форм жизни — бактерий, архей и эукариот — неродственны друг другу. Как же так получилось, что такой важнейший для любой клетки процесс как репликация ДНК у трех доменов жизни катализируют совершенно разные белки? Недавно группа ученых во главе с Евгением Куниным (Eugene V. Koonin) предложила новый изящный сценарий возможного происхождения репликации ДНК. Но прежде чем приступить к его обсуждению, давайте рассмотрим, какие вообще бывают ДНК-полимеразы.
ДНК-полимеразы бактерий, архей и эукариот относятся к трем разным белковым семействам и различаются друг от друга, прежде всего, организацией активного центра, который и катализирует присоединение новых нуклеотидов к растущей комплементарной цепи ДНК.
У эукариот, архей из типа Crenarchaeota и многих крупных ДНК-содержащих вирусов основная ДНК-полимераза, задействованная в репликации, относится к семейству полимераз B (PolB). Полимеразы из семейства PolB есть у некоторых бактерий и архей, не относящихся к Crenarchaeota, но у них она участвует не в репликации, а в репарации, восстанавливая бреши в поврежденных цепях ДНК. Ферменты PolB объединяет одинаковое устройство каталитического домена: его основу составляет мотив под названием RRM-ладонь (RNA recognition motif) (рис. 1, B). «Ладонью» (англ. palm) принято называть домен ДНК-полимеразы, который катализирует присоединение нового нуклеотида к цепочке (домен под названием «палец» (finger) обеспечивает образование комплементарных связей между новоприсоединенным доменом и матричной цепью, а домен «большой палец» (thumb) нужен для эффективного перемещения фермента по матричной цепи; подробнее см., например, T. A. Steitz, 1999. DNA Polymerases: Structural Diversity and Common Mechanisms). Домены получили свои говорящие названия из-за сходства их расположения друг относительно друга с расположением пальцев и ладони кисти руки.
Примечательно, что очень похожую структуру с мотивом RRM имеет каталитический центр ферментов из числа обратных транскриптаз, которые синтезируют РНК на матрице ДНК, и вирусных РНК-зависимых РНК-полимераз, которые синтезируют РНК на матрице РНК. Кроме того, активный центр на основе мотива RRM-ладонь имеют праймазы — ферменты, синтезирующие РНК-затравки в начале удвоения ДНК у архей, эукариот и вирусов, геном которых представлен двуцепочечной ДНК. Производное мотива RRM-ладонь имеется у полимераз семейства PolA. Эти полимеразы задействованы в репаративном синтезе ДНК у бактерий и являются главным репликативным ферментом у некоторых фагов, а также в митохондриях растений и грибов.
У архей, за исключением представителей типа Crenarchaeota и некоторых других видов, основная репликативная ДНК-полимераза относится к семейству полимераз D (PolD). Каталитический центр PolD образован двумя структурными мотивами, известными как двойной ψβ-бочонок (DPBB, от англ. Double Psi Beta Barrel). Два мотива DPBB составляют основу активного центра основного фермента транскрипции — РНК-полимеразы — у большинства форм жизни: бактерий, архей, многих ДНК-вирусов (рис. 1, A).
Бактериальная репликативная ДНК-полимераза принадлежит к семейству полимераз C (PolC). Каталитический центр имеет PolC иную структуру, чем PolB. Его основу составляет структурный мотив, известный как Polβ-подобная нуклеотидилтрансфераза (рис. 1, C). За пределами домена бактерий PolC практически не известны: исключение составляют несколько плохо изученных бактериофагов. Редкость полимераз PolC среди фагов, вероятно, свидетельствует о том, что PolC попали к вирусам от бактерий относительно недавно и не успели широко распространиться.
Как вы, наверное, уже заметили, различные семейства ДНК-полимераз существенно ближе к некоторым РНК-полимеразам, чем друг к другу. Долгое время ученые полагали, что репликация ДНК в ходе эволюции возникала несколько раз. Некоторые даже придерживались мнения, что геном LUCA был представлен молекулой РНК, поэтому и механизм репликации ДНК организмы из разных доменов «изобретали» независимо друг от друга.
В последние годы благодаря развитию метагеномики, позволяющей получать последовательности геномов микроорганизмов и вирусов прямо из образцов окружающей среды без культивации в лаборатории, базы данных пополнились последовательностями ДНК-полимераз множества новых организмов. Теперь уже стало возможным проследить определенные закономерности эволюции ДНК-полимераз, сравнивая последовательности миллионов генов, кодирующих ДНК-полимеразы у самых разных организмов. Кроме того, многие ДНК-полимеразы микроорганизмов и вирусов ученые сумели закристаллизовать и далее получить их пространственные структуры. Ученые из группы Кунина проанализировали новые данные о сходстве ДНК- и РНК-полимераз различных организмов и на их основании разработали вероятный сценарий возникновения и эволюции репликации ДНК.
Исследователи полагают, что наличие РНК-полимераз с двумя мотивами DPBB (являющихся основными ферментами транскрипции) у всех доменов жизни свидетельствует о том, что РНК-полимераза такого же устройства имелась еще у LUCA. Примечательно, что, хотя современные РНК-полимеразы с двумя мотивами DPBB обычно функционируют как ДНК-зависимые ферменты, при некоторых условиях они могут синтезировать РНК на матрице РНК. Например, растительные инфекционные агенты вироиды, имеющие мелкие РНК-геномы, заставляют РНК-полимеразу II растений функционировать как РНК-зависимую РНК-полимеразу, хотя как таковые РНК-зависимые РНК-полимеразы имеются только у РНК-содержащих вирусов. Ученые заключили, что предковая полимераза, содержащая мотив DPBB, появилась еще до того, как ДНК стала основной формой хранения генетической информации, и была РНК-зависимой РНК-полимеразой. Сам мотив DPBB, вероятно, первоначально служил вспомогательным, лишенным каталитической активности РНК-связывающим доменом, а роль РНК-зависимой РНК-полимеразы выполнял рибозим. Однако белок, содержащий не один, а два мотива DPBB в результате удвоения соответствующего участка гена, уже сам обладал полимеразной активностью и постепенно вытеснил менее эффективный рибозим, став настоящей РНК-зависимой РНК-полимеразой.
Появление клеток, геном которых представлен ДНК, сопровождалось разделением древней РНК-зависимой РНК-полимеразы с двумя мотивами DPBB на две эволюционные ветви: первая ветвь была родственна современным PolD архей и включала ДНК-зависимые ДНК-полимеразы, а вторую ветвь образовались РНК-полимеразы, «научившиеся» использовать ДНК в качестве матрицы и ставшие первыми ферментами, осуществляющими транскрипцию. Появление репликативных ДНК-зависимых ДНК-полимераз сопровождалось присоединением к DPBB-полимеразе нескольких дополнительных доменов, облегчающих взаимодействие с ДНК, в частности, доменов, содержащих ДНК-связывающие мотивы типа цинковые пальцы. Авторы гипотезы полагают, что у LUCA имелась репликационная ДНК-полимераза именно такого типа (то есть содержащая мотив DPBB и входящая в семейство PolD), а из современных организмов ее сохранили большинство архей.
Когда уже после LUCA произошло разделение клеточной жизни на архей и бактерий (эукариоты, по современным представлениям, являются обособленной группой архей, см. Новооткрытый микроб заполняет брешь между прокариотами и эукариотами, «Элементы», 12.05.2015), у бактерий DPBB-содержащая ДНК-полимераза была вытеснена ДНК-полимеразой семейства PolC, которая произошла от древней нуклеотидилтрансферазы семейства Polβ.
Поскольку мотив RRM очень консервативный и древний (он появился, вероятно, еще в эпоху РНК-мира), полимеразы семейства PolB, основу каталитического центра которых составляет RRM, возможно, появились у мобильных генетических элементов и вирусов, паразитировавших на древних протоклетках еще до возникновения LUCA. В частности, возможным предком PolB является обратная транскриптаза первых ретротранспозонов. У архей ДНК-полимеразы семейства PolB появлялись несколько раз, вероятно, благодаря заимствованиям у вирусов, у которых ферменты этой группы широко распространены. Появление полимераз PolB у бактерий также, по всей видимости, связано с заимствованием этого фермента у вирусов. У эукариот полимеразы PolB тоже появились благодаря вирусам, причем впоследствии эукариотические ДНК-полимеразы PolD, сохранившиеся от предков-архей, были полностью вытеснены PolB.
Происхождение полимераз PolA, содержащих мотив RRM и отличающихся высокой консервативностью, вызывает много вопросов. Возможно, PolA впервые появились у вирусов из предковой RRM-полимеразы, а затем были перенесены бактериям. Примечательно, что подобный сценарий был реализован у митохондрий, у которых PolA служит главным репликативным ферментом: вероятнее всего, митохондрии получили PolA от фагов. Таким образом, заимствование клеткой вирусных ферментов, которые по эффективности зачастую превосходят клеточные аналоги, происходило несколько раз в ходе эволюции репликации ДНК.
Таким образом, согласно новой гипотезе Кунина и соавторов, современные ДНК- и РНК-полимеразы произошли от общего белка-предка, который функционировал как РНК-зависимая РНК-полимераза. Существующие возможности молекулярной биологии позволяют, хотя и с большим трудом, реконструировать древние формы РНК- и ДНК-полимераз. И, хотя изложенный выше сценарий в значительной степени остается гипотетическим, реконструкция предковых полимераз может позволить проверить его экспериментально.
Источник: elementy.ru





