













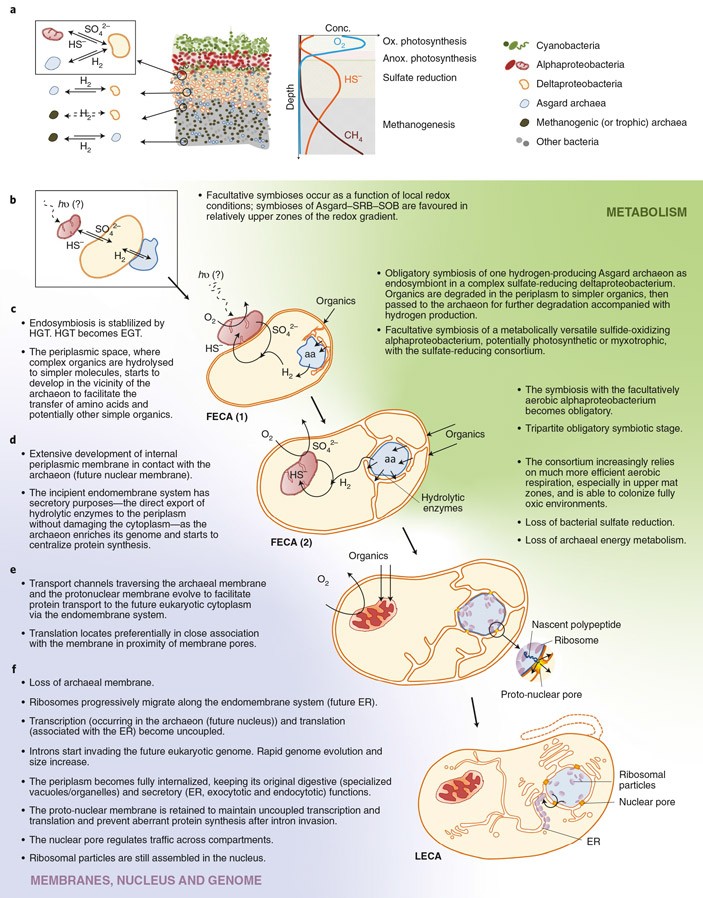


Рис. 1. Обновленная синтрофная гипотеза. a — общая структура сообщества цианобактериального мата, где предположительно началось становление эукариот. b — стадия факультативного симбиоза трех партнеров: гетеротрофной асгардархеи, сульфатредуцирующей дельта-протеобактерии и альфа-протеобактерии, окисляющей сероводород (возможно, она была еще и фотосинтезирующей). c — стабилизация симбиоза за счет горизонтального переноса генов и переход его в эндосимбиоз, когда один из партнеров поселяется внутри другого, в данном случае — архея внутри дельта-протеобактерии. d — симбиоз с альфа-протеобактерией тоже принимает характер эндосимбиоза, одновременно у хозяина (то есть у дельта-протеобактерии) развивается система внутренних мембран. e — дельта-протеобактерия отказывается от восстановления сульфата и переходит к брожению, альфа-протеобактерия отказывается от окисления сульфидов и переходит к кислородному дыханию, а архея вообще навсегда теряет собственный энергетический обмен; кроме того, начинают формироваться ядерные поры. f — исчезновение архейной мембраны и окончательное формирование ядерных пор, приводящее к свойственному эукариотам разобщению процессов транскрипции и трансляции; на этой стадии перед нами полностью сформированная эукариотная клетка. Иллюстрация из обсуждаемой статьи в Nature Microbiology
Известные испанские (работающие во Франции) микробиологи Пурификасьон Лопес-Гарсия и Давид Морейра предложили новую гипотезу происхождения эукариот, которая называется обновленной синтрофной гипотезой или гипотезой HS-синтрофии. Она учитывает данные, полученные в результате прошлогоднего открытия живого представителя асгардархей. Согласно гипотезе HS-синтрофии, возникновение эукариот было результатом метаболического симбиоза между тремя партнерами: асгардархеей, дельта-протеобактерией и альфа-протеобактерией, при этом на первых стадиях симбиоза имел большое значение обмен серы. В конце концов асгардархея образовала содержимое ядра, дельта-протеобактерия — основу цитоплазмы, а альфа-протеобактерия — митохондрии.
Альтернативы симбиоза
Со времен совершённых в 1970-х годах великих молекулярно-биологических открытий Карла Вёзе (Carl Richard Woese; см. Эволюция по Вёзе) биологи стали делить все современные живые существа (кроме вирусов) на бактерий, архей и эукариот. К эукариотам относятся самые крупные и сложные организмы на Земле: многоклеточные животные, шляпочные грибы, бурые водоросли, высшие растения. Разумеется, эукариоты состоят из клеток, но эукариотная клетка негомологична клеткам бактерий и архей. Это гораздо более сложная структура, возникшая в результате объединения нескольких (как минимум двух) «элементарных клеток» и последующей пересборки их компонентов.
Представления о том, как именно это произошло, сильно менялись по мере накопления новых сведений. Лет тридцать назад в биологии утвердилась классическая симбиогенетическая теория, связанная с именем Линн Маргулис (Lynn Margulis; см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). Предполагалось, что протоэукариоты, которые, вероятно, были сестринской группой по отношению к археям, сначала самостоятельно приобрели большинство эукариотных признаков — сложный подвижный цитоскелет, внутреннюю систему мембранных полостей, ядро и фагоцитоз, — а потом захватили и поселили у себя внутри альфа-протеобактерий, постепенно превратившихся в митохондрии.
Однако чем дальше, тем больше появлялось фактов, противоречащих этой почтенной «маргулисовской» модели. Во-первых, эукариот, никогда не имевших митохондрий, по-видимому, не существует (см. Обнаружены одноклеточные организмы с ядром, но без митохондрий, «Элементы», 18.05.2016). Во-вторых, не подтверждается, что фагоцитоз появился раньше, чем митохондрии: как раз наоборот, многое указывает на то, что до союза с митохондриями фагоцитоз был бы невозможен (W. F. Martin et al., 2017. The physiology of phagocytosis in the context of mitochondrial origin). В-третьих, молекулярная систематика свидетельствует, что архейный предок эукариот был не таинственной и абстрактной «сестринской группой архей», а просто археей (см. Описан новый надтип архей, к которому относятся предки эукариот, «Элементы», 16.01.2017). И в-четвертых, недавно открытый современный представитель архей, близко родственный эукариотам, не проявляет никаких признаков переходного состояния: ни подвижного цитоскелета, ни внутренних мембранных полостей, ни ядра, ни фагоцитоза у него нет (см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019).
Все эти данные настолько серьезно изменили представления о происхождении эукариот, что в этой области теперь вполне можно говорить о смене парадигм. Как известно, само понятие «парадигма» предполагает существование семейства гипотез, которые основаны на общих предпосылках, но могут — и даже должны — конкурировать друг с другом и противоречить друг другу в отдельных утверждениях. Новая парадигма происхождения эукариот — не исключение. Она начала формироваться больше двадцати лет назад, когда фактов, подтверждающих новые взгляды, было еще немного, и уже тогда в ее рамках обсуждались разные эволюционные сценарии.
В 1998 году Билл Мартин (William Martin) и Миклош Мюллер (Miklós Müller) опубликовали водородную гипотезу происхождения эукариот (W. Martin, M. Müller, 1998. The hydrogen hypothesis for the first eukaryote). Они предположили, что эукариоты возникли в результате тесной связи между двумя микроорганизмами: альфа-протеобактерией, которая питалась путем брожения, и метаногенной археей. Альфа-протеобактерия выделяла конечные продукты брожения, в том числе углекислоту и водород (см. Гидрогеносомы). А метаногенная архея, которая живет именно за счет восстановления углекислоты с помощью водорода, с радостью эти продукты захватывала. Ключевым фактором при этом был водород, который относительно редко встречается на Земле, но зато легко передается от клетки к клетке, потому что его молекулы очень маленькие. В свою очередь, метаногенная архея делилась с альфа-протеобактерией восстановленными органическими молекулами, которыми бродильщик может питаться. Для метаногенной археи этот союз был очень выгоден, поэтому она со всех сторон окружила альфа-протеобактерию своими выростами, чтобы увеличить поверхность контакта между клетками. В дальнейшем альфа-протеобактерия переключилась с брожения на кислородное дыхание (подобные способности есть у многих членов этого класса) и стала митохондрией. А метаболизм бывшей метаногенной археи перестроился таким образом, чтобы самостоятельно проводить частичное окисление восстановленных органических молекул, захваченных теперь уже из внешней среды, и передавать промежуточные продукты для окончательного окисления в митохондрию, получая взамен АТФ. Таким образом бывшая архея перестала нуждаться в водороде и заодно превратилась из автотрофного организма в гетеротрофный. Она-то, в соответствии с водородной гипотезой, и стала основой эукариотной клетки, образование которой, как видим, без участия предков митохондрий было бы немыслимо.
В том же 1998 году Давид Морейра (David Moreira) и Пурификасьон Лопес-Гарсия (Purificación López-García) опубликовали синтрофную гипотезу происхождения эукариот (D. Moreira, P. Lopez-Garcia, 1998. Symbiosis between methanogenic archaea and δ-proteobacteria as the origin of eukaryotes: the syntrophic hypothesis). Для Морейры и Лопес-Гарсии концепция Мартина и Мюллера была слишком простой. По их мнению, тесную связь с метаногенной археей образовала не альфа-протеобактерия, а дельта-протеобактерия: представители этого класса вступают в такие симбиозы и сейчас, и нет причин считать, что они не делали то же самое в древние времена. Разлагая органические молекулы, дельта-протеобактерия превращала их в углекислоту и водород, которыми щедро делилась с метаногенной археей. Последняя образовала для увеличения поверхности контакта выросты, между которыми многочисленные дельта-протеобактерии стали селиться. Потом эти дельта-протеобактерии слились друг с другом, потеряли генетический аппарат и стали цитоплазмой эукариотной клетки. А бывшая клетка метаногенной археи, которая свой генетический аппарат сохранила, стала ядром. До этого момента все участники симбиоза были анаэробными, то есть не использовали кислород в обмене веществ. Аэробной эукариотная клетка стала, только приобретя еще одного партнера — альфа-протеобактерию, будущую митохондрию.
Как видим, гипотеза Лопес-Гарсии и Морейры предполагает, что партнеров, объединившихся в эукариотную клетку, было не два, а три. Проблема этой гипотезы в том, что она предсказывает решающий вклад дельта-протеобактерий в геном эукариот, которого на самом деле не наблюдается. У эукариот, правда, есть гены дельта-протеобактериального происхождения, но у них есть и гены, заимствованные еще от нескольких групп бактерий: альфа-протеобактерий, гамма-протеобактерий, грамположительных бактерий и других (см. E. V. Koonin, 2010. The origin and early evolution of eukaryotes in the light of phylogenomics). Никак нельзя сказать, что дельта-протеобактерии на этом фоне вне конкуренции. Но в любом случае эти данные свидетельствуют о том, что формирование эукариот происходило в очень плотном многовидовом архейно-бактериальном сообществе, где были все условия не только для симбиоза, но и для обмена генами (рис. 2). В этом Лопес-Гарсия и Морейра, безусловно, правы.

Рис. 2. Водородная и синтрофная гипотезы. Сверху изображено исходное многовидовое сообщество. Ацетат, наряду с углекислотой, является как одним из продуктов брожения, так и одним из исходных реагентов метаногенеза. На схеме синтрофной гипотезы отражена идея авторов, что альфа-протеобактерия сначала была метанотрофной: она питалась метаном, который выделяла метаногенная архея, и превращала его в углекислоту, тем самым замыкая еще один контур симбиотического взаимодействия. На этой стадии все партнеры были анаэробными. Освоение аэробных условий произошло позже, после переключения альфа-протеобактерии на кислородный метаболизм. Все остальные пояснения — в тексте. Иллюстрация из статьи P. López-Garcı́a, D. Moreira, 1999. Metabolic symbiosis at the origin of eukaryotes
Кроме того, среди дельта-протеобактерий много сульфатредукторов, которые в процессе получения энергии превращают сульфаты в сульфиды. Как известно, в состав этих молекул входит сера. Большинство всех сульфатредуцирующих микроорганизмов относится именно к дельта-протеобактериям. Лопес-Гарсия и Морейра, конечно, знали это, но в первоначальной версии их гипотезы обмен серы не сыграл никакой роли — эта деталь осталась «лишней». Тем не менее они сразу же отметили, что в современной природе самый распространенный вариант архейно-бактериального симбиоза — это симбиоз метаногенных архей с бактериями-сульфатредукторами (P. López-Garcı́a, D. Moreira, 1999. Metabolic symbiosis at the origin of eukaryotes). Запомним эту подробность.
Синтрофная парадигма
Водородная и синтрофная гипотезы разрабатывались совершенно независимо (до выхода публикаций их авторы ничего не знали об идеях друг друга), но оказались поразительно похожими. Они сходятся как минимум в четырех пунктах:
1) союз предков эукариот возник не через пожирание одного партнера другим (фагоцитоз), а через мирное сожительство, сопровождавшееся обменом продуктами метаболизма (синтрофия);
2) ключевым метаболитом, на котором этот союз в первую очередь держался, был водород;
3) в ходе развития симбиоза один партнер охватывал другого своими выростами;
4) среди партнеров была метаногенная архея.
После открытия реальных современных архейных родственников эукариот все четыре тезиса подтвердились — но подтвердились неожиданным образом (H. Imachi et al., 2020. Isolation of an archaeon at the prokaryote–eukaryote interface). Можно предположить, что при виде этих данных авторы и водородной, и синтрофной гипотез пережили крайне противоречивые чувства. С одной стороны, в природе найдено реальное синтрофное сожительство трех партнеров, два из которых — это… метаногенная архея и сульфатредуцирующая дельта-протеобактерия! С другой же стороны, родичи эукариот там представлены вовсе не метаногенной археей (близкое родство этой группы с эукариотами вообще не подтверждается), а третьим партнером — локиархеей. И вот она-то преподнесла нашим авторам сюрпризы. Во-первых, она оказалась не автотрофным метаногеном, а гетеротрофным бродильщиком, питающимся аминокислотами. И во-вторых, она оказалась не акцептором водорода, а его донором (при таком типе метаболизма иначе и быть не может). Как в старом анекдоте: не в покер, а на скачках, и не выиграл, а проиграл, а в остальном все верно.
В такой сложной ситуации бывает очень полезно, условно говоря, перейти на один уровень вверх, чтобы охватить пространство логических возможностей как можно полнее (рис. 3). Материала для этого хватает: в последние два десятилетия наука, мягко говоря, не стояла на месте. В обзоре, вышедшем в Nature в апреле 2020 года, уже знакомые нам Пурификасьон Лопес-Гарсия и Давид Морейра насчитывают минимум семь гипотез происхождения эукариот, основанных на синтрофии или метаболическом симбиозе (это одно и то же).

Рис. 3. Краткая сводка гипотез метаболического симбиоза. Иллюстрация из обсуждаемой статьи в Nature Microbiology
1. Гипотеза серной синтрофии (D. Searcy, 2003. Metabolic integration during the evolutionary origin of mitochondria). Предполагается, что предки митохондрий окисляли сероводород до молекулярной серы, которую архейный партнер использовал в своем метаболизме и восстанавливал обратно до сероводорода. Эта гипотеза основана в первую очередь на том, что современные свободноживущие родственники митохондрий — пурпурные альфа-протеобактерии — действительно умеют окислять сероводород. Но вот идея, что архейный предок эукариот специализировался на восстановлении серы, имеет гораздо меньше подтверждений. В любом случае эта гипотеза интересна тем, что первой привлекла внимание к обмену серы — элемента, который необыкновенно удобен для всевозможных окислительно-восстановительных реакций и был доступен в изобилии на древней Земле.
2. Классическая водородная гипотеза Мартина и Мюллера, о которой мы уже говорили.
3. Обновленная версия водородной гипотезы, предложенная после открытия локиархей (F. Sousa et al., 2016. Lokiarchaeon is hydrogen dependent). Она принципиально не отличалась от классической версии, но в ней было добавлено основанное на новых данных уточнение: архейный предок эукариот превращал углекислоту и водород в органические молекулы с помощью древнейшего биохимического пути Вуда — Льюнгдаля (см. Источником энергии для древнейшей жизни мог служить ацетилфосфат, «Элементы», 04.07.2018). Разумеется, такой предок должен был бы быть водородозависимым.
4. Гипотеза обратного потока, предложенная в 2019 году в результате детальных исследований геномов локиархей, и шире — асгардархей (Asgardarchaeota, см. A. Spang et al., 2019. Proposal of the reverse flow model for the origin of the eukaryotic cell based on comparative analyses of Asgard archaeal metabolism). Здесь впервые была высказана идея, что водород переносился не в ту сторону. Согласно гипотезе обратного потока, архейный предок архей был не автотрофом (как в водородной и синтрофной гипотезах), а гетеротрофом, разлагавшим сложную органику путем брожения. В результате он получал, во-первых, мелкие органические молекулы, а во-вторых, водород, которым мог пользоваться любой желающий — в том числе и предки митохондрий, если они умели использовать его в метаболизме (а это не исключено; см. Hydrogen oxidizing bacteria). Таким образом, архейный предок эукариот был не акцептором, а донором водорода. Это предсказание, основанное на тот момент только на чтении обрывков геномов, получило блестящее подтверждение через четыре месяца, когда были описаны живые локиархеи.
5. Гипотеза «трех E», предложенная теми самыми японскими авторами, которые открыли живых локиархей (см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019). Согласно этой гипотезе, архейный предок эукариот был бродильщиком и питался аминокислотами. В результате брожения получались мелкие органические молекулы и водород. Органические побочные продукты «главный герой» отдавал альфа-протеобактериям, а водород — метаногенным археям и (или) сульфатредуцирующим дельта-протеобактериям, которые с удовольствием использовали его, соответственно, для восстановления углекислоты или сульфатов. После Великого кислородного события (Great Oxidation Event) океан обогатился сульфатами, и питавшиеся ими сульфатредукторы вытеснили метаногенных архей из симбиоза. Осталось три участника: асгардархея (бродильщик), альфа-протеобактерия («сжигатель» продуктов брожения) и дельта-протеобактерия (сульфатредуктор). Со временем в окружающей среде становилось все больше кислорода, и это повышало ценность альфа-протеобактерий, умеющих его утилизировать. Поэтому архейный партнер сначала запутал альфа-протеобактерий в своих выростах (entangled), потом окружил ими со всех сторон (engulfed), а потом полностью замкнул внутри своей клетки — эндогенизировал (endogenized). Так возникли митохондрии. Что же касается дельта-протеобактерий, то их участие в итоге ограничилось передачей будущим эукариотам некоторых генов. Если у кого-то из читателей от сложности этого сценария голова пойдет кругом, ему можно посочувствовать, но похоже, что никакие более простые объяснения тут не сработают: реальные эволюционные события были еще сложнее. Впрочем, это не обязательно означает, что гипотеза «трех E» во всем верна.
6. Уже известная нам классическая синтрофная гипотеза Лопес-Гарсии и Морейры. Сейчас авторы называют ее HM-синтрофией: H — водород, M — метан.
7. Обновленная версия синтрофной гипотезы, которую Лопес-Гарсия и Морейра предлагают в обсуждаемой статье. Эту версию они называют HS-синтрофией: H — водород, S — сера. Она опубликована впервые, и о ней мы должны поговорить подробнее (см. рис. 1).
HS-гипотеза
Альфа-протеобактерии, — к которым, еще раз напомним, относятся митохондрии, — известны как микроорганизмы с очень широкими метаболическими возможностями. Нет ровным счетом ничего удивительного в предположении, что непосредственный предок митохондрий наряду с дыханием (а может, и вместо него) владел способностью окислять сероводород до молекулярной серы или до сульфатов. Как мы знаем, именно распространенность у современных альфа-протеобактерий такого типа метаболизма послужила основанием для гипотезы серной синтрофии. Но большой популярности она не получила: не было надежных подтверждений, что второй симбионт (архейный) мог бы включиться в этот серный обмен.
Теперь, однако, у нас есть свидетельства вхождения в дело еще одного симбионта — сульфатредуцирующей дельта-протеобактерии. Для серной синтрофии это идеальный вариант. Альфа-протеобактерия окисляет сульфиды до сульфатов, а дельта-протеобактерия питается сульфатами, превращая их вновь в сульфиды. Оба партнера дают друг другу самое нужное.
Однако для того, чтобы превращать сульфаты в сульфиды, дельта-протеобактерии необходим еще и водород. Где она его возьмет? Правильно: у третьего партнера — гетеротрофной археи, у которой водород является побочным продуктом брожения, бесполезным для нее самой, но очень полезным для привлечения симбионтов.
В свою очередь, дельта-протеобактерия может поставлять архее питательную органику, которую та будет сбраживать, получая водород и мелкие органические молекулы. Водород пойдет на поддержку дельта-протеобактерии, а через нее и альфа-протеобактерии. Ну и «бонусом» альфа-протеобактерия может попытаться захватить органические продукты брожения, чтобы окончательно разложить их с участием кислорода, получив энергию. Цепочка из трех звеньев замыкается.
Лопес-Гарсия и Морейра не отвергают окончательно свою старую HM-гипотезу: они справедливо замечают, что метаногенез — древнейший тип метаболизма, скорее всего исходный для архей, а может быть, и вообще для всех клеточных организмов (см. Источником энергии для древнейшей жизни мог служить ацетилфосфат, «Элементы», 04.07.2018). Исключать, что архейный предок эукариот мог быть метаногеном, нельзя по той простой причине, что чей угодно предок мог когда-нибудь быть метаногеном. Но факты, известные на данный момент, скорее свидетельствуют в пользу только что изложенной HS-гипотезы. Тут все фрагменты пазла более-менее встают на места.
Лопес-Гарсия и Морейра вновь и вновь обращают внимание на несколько фактов, которые хорошо известны уже лет двадцать. Несмотря на то, что эукариоты формально считаются потомками архей, больше половины их генов имеет бактериальное происхождение. От архей эукариоты получили преимущественно «информационные» гены, продукты которых формируют наследственный аппарат, а от бактерий — преимущественно «операционные» гены, продукты которых управляют метаболизмом, энергетикой, транспортом, сигнализацией и синтезом мембран (см. Подсчитан вклад архейных и бактериальных генов в эукариотные геномы, «Элементы», 07.04.2020). Если говорить совсем примитивно, эукариотная клетка выглядит так, будто содержимое ядра она унаследовала исключительно от архей, а все остальное — в основном от бактерий. Ну так почему бы и нет? — спрашивают Лопес-Гарсия и Морейра. Дельта-протеобактерии, кроме всего прочего, известны своими крупными размерами и склонностью к образованию колоний (см. Миксобактерии). Так что гипотеза, согласно которой дельта-протеобактерия (или группа таких бактерий) охватила архейного симбионта и в конце концов поселила его у себя внутри, остается в силе. А мембранная система, внутри которой архейный симбионт в итоге оказался, послужила основой для эндоплазматической сети, ядра, лизосом, пероксисом, аппарата Гольджи, пищеварительных вакуолей и прочей мембранной «начинки», которой набита любая уважающая себя эукариотная клетка.
Ядро, по этой гипотезе, сначала представляло собой систему впячиваний мембран дельта-протеобактерии, окружавших архейного симбионта. После появления ядерных пор, служащих для контролируемого транспорта молекул из ядра в цитоплазму и обратно, связь ядра с внешней мембраной исчезла. Мембрана археи тоже исчезла за ненадобностью, как и архейные энергетические системы: все эти функции взял на себя бактериальный компонент. От археи остался голый геном с аппаратом синтеза белка.
Если HS-гипотеза верна, то на первом этапе сообщество предков эукариот вполне могло быть полностью анаэробным. Нельзя исключить, что оно сложилось задолго до Великого кислородного события. Переход к аэробному образу жизни произошел гораздо позже, за счет переключения альфа-протеобактериального партнера с окисления сульфидов на кислородное дыхание. Это — типичная смена функций, аналогичная той, которая той, которая лежит в основе возникновения новых сложных адаптаций, например, у животных. Именно смена функций обеспечивает постепенность эволюции, даже если начальное и конечное звенья эволюционной последовательности выглядят совершенно непохожими друг на друга.
Тут надо учитывать, что на синтрофные отношения (в отличие от фагоцитоза) не распространяется принцип «все или ничего». У хищника выбор невелик: он может либо съесть жертву, либо не съесть. Но синтрофные отношения, где никто никого не ест и стороны мирно обмениваются молекулами, почти как субъекты рынка товарами, допускают гораздо больше градаций. Эти отношения могут быть более сильными или менее сильными, могут по ходу дела включать или исключать дополнительных участников и тому подобное. Рискнем сказать, что синтрофные отношения по своей природе избыточны. Несомненно, это облегчает смену как партнеров, так и ролей, которые играют эти партнеры, и в целом дает много возможностей для эволюционного «маневрирования».
У новой гипотезы Лопес-Гарсии и Морейры, несомненно, есть сильные стороны. Во-первых, она удачно совмещает две трудносовместимые вещи: бесспорно колоссальный вклад бактерий в эукариотный геном и позднемитохондриальный сценарий (см. Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Существуют биоинформатические данные, согласно которым митохондрии эволюционно моложе, чем любые другие важные части эукариотной клетки (эндоплазматическая сеть, аппарат Гольджи и тому подобное). Но если митохондрии возникли поздно, то когда же они успели так сильно повлиять на всю внутриклеточную архитектуру? Возможный ответ состоит в том, что дело не в митохондриях — точнее, не только в них. Они просто вселились в ту же самую дельта-протеобактерию, которая чуть раньше стала приютом для археи. А может, и в протоэукариотную клетку, когда та была уже в основных чертах «смонтирована» из этих двух источников. Кстати, из упомянутых биоинформатических данных вроде как следует, что именно дельта-протеобактериальные гены занимают в эукариотном геноме следующее место после альфа-протеобактериальных (если не считать архейных; A. A. Pittis, T. Gabaldon, 2016. Late acquisition of mitochondria by a host with chimaeric prokaryotic ancestry).
Во-вторых, HS-гипотеза (как и HM-гипотеза) снимает проблему преемственности мембран. Вкратце эта проблема состоит в следующем. Архейные и бактериальные клеточные мембраны состоят из совершенно различных молекул, принадлежащих к разным классам органических веществ и отличающихся на уровне оптической изомерии, при этом у эукариот все мембраны относятся к бактериальному типу. Внутри одной мембраны архейные и бактериальные составляющие плохо совместимы: правда, пару лет назад генно-инженерными методами удалось получить «гибридную» кишечную палочку, у которой до 30% мембранных липидов были архейными, но дальнейшее увеличение их доли делает клетку нежизнеспособной (A. Caforio et al., 2018. Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane). Так что вообразить устойчивое переходное состояние здесь трудно. Если эукариотная клетка непосредственно произошла от архейной, то в ней должно было произойти очень быстрое тотальное замещение всех мембран — вероятно, с помощью альфа-протеобактерий, ставших митохондриями. Гипотезы, объясняющие этот механизм, существуют, и они по-своему убедительны (см. S. B. Gould et al., 2016. Bacterial vesicle secretion and the evolutionary origin of the eukaryotic endomembrane system и краткий обзор по этой работе). Но в рамках HS-гипотезы никаких механизмов замещения придумывать не надо: она предполагает, что мембраны были напрямую унаследованы эукариотами от бактерий, вот и вся загадка. Такое решение выглядит подозрительно простым, учитывая, что эукариотизация явно была процессом в высшей степени динамическим, но это ни в коем случае нельзя считать возражением по существу. Посмотрим, что скажут по этому поводу другие специалисты и, главное, что обнаружат исследования современных асгардархей: тут наверняка будут новые сюрпризы. До полного раскрытия тайны происхождения эукариот пока еще далеко. Чем больше мы узнаём, тем больше возникает новых вопросов, в том числе таких, которые еще недавно было невозможно даже задать. Здесь предельно наглядно проявляет себя принцип Джона Арчибальда Уилера (John Archibald Wheeler): «По мере того, как растет остров знания, растет и береговая линия соприкосновения с неведомым».
Источник: elementy.ru





