













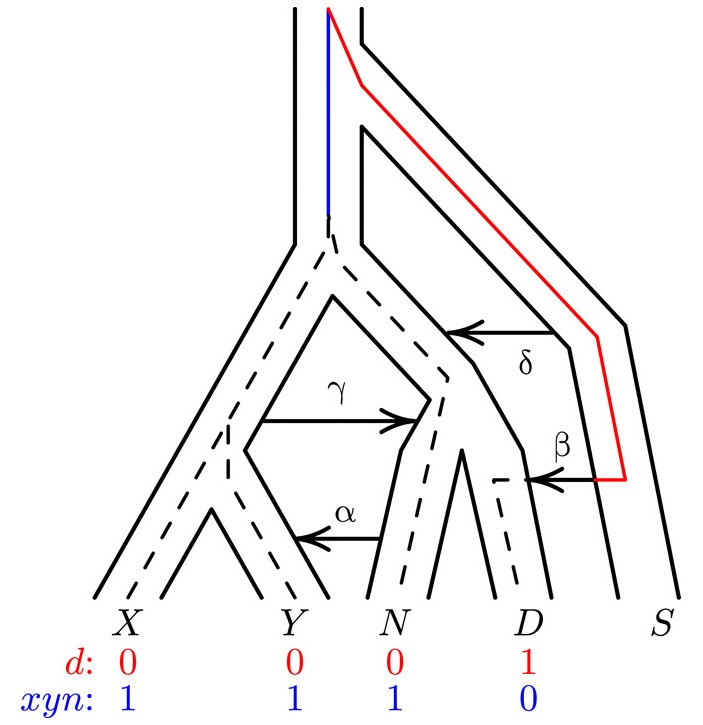

Рис. 1. Реконструированная генеалогия пяти человеческих популяций: X — современные африканцы, Y — современные европейцы, N — неандертальцы, D — денисовцы, S — неизвестная сверхархаичная популяция (Superarchaics). Схема также иллюстрирует идею о том, что частота встречаемости разных комбинаций примитивных и продвинутых аллелей (nucleotide site patterns) несет информацию об истории изучаемых популяций. Нулями обозначены примитивные аллели, единицами — производные. Стрелки с греческими буквами — эпизоды привноса чужих генов в результате гибридизации. Синими, красными и пунктирными линиями показана генеалогия одного конкретного нуклеотидного сайта. При такой генеалогии мутация, произошедшая на красной ветви, порождает комбинацию примитивных и производных аллелей d (0001), а мутация, произошедшая на синей ветви — комбинацию xyn (1110). Прочие пояснения в тексте. Рисунок из обсуждаемой статьи в Science Advances
Новый метод анализа геномных данных позволил американским ученым обосновать идею о том, что общие предки неандертальцев и денисовцев («неандерсовцы»), выйдя из Африки, скрещивались с некой «сверхархаичной» человеческой популяцией, обособившейся около 2 млн лет назад. Скорее всего, это были потомки самых первых выходцев из Африки — ранних эректусов, древнейшие следы присутствия которых в Евразии имеют возраст около 1,8 млн лет (люди из Дманиси). Позднее денисовцы, по-видимому, снова скрещивались с представителями этой архаичной популяции, но другой ее ветви. Наряду с новыми данными о присутствии в геномах современных африканских народов различных архаичных примесей, открытие показало, что отдаленная гибридизация периодически происходила на протяжении чуть ли не всей истории человеческого рода.
Современное человечество генетически довольно однородно. Все мы — от европейцев и восточноазиатов до бушменов и коренных австралийцев — вне всяких сомнений принадлежим к одному виду и по большей части происходим от одной и той же предковой популяции ранних сапиенсов, которая жила в Африке примерно 200–300 тысяч лет назад.
Но такая гомогенность не всегда была присуща человеческому роду. Это относительно новое явление, почти наверняка связанное с тем, что наши прямые предки просто-напросто вытеснили все остальные ветви рода Homo. Еще совсем недавно, каких-то 50–70 тысяч лет назад, на Земле одновременно проживало несколько сильно различающихся человеческих групп: сапиенсы, неандертальцы, денисовцы, островные «хоббиты» с Флореса и Лусона и, возможно, кто-то еще. Многие из этих групп, скорее всего, заслуживают статуса самостоятельных видов, потому что успели накопить достаточно генетических различий, чтобы гибридное потомство имело пониженную приспособленность (это называют частичной постзиготической репродуктивной изоляцией; см.: Между сапиенсами и неандертальцами существовала частичная репродуктивная изоляция, «Элементы», 03.02.2014). Впрочем, это не мешало им при случае всё-таки скрещиваться, производя это самое гибридное потомство и таким образом подмешивая свои гены в генофонды дальних родственников.
Благодаря быстрому развитию палеогенетики и сравнительной геномики постоянно выясняются новые подробности о многочисленных (как теперь уже ясно) эпизодах гибридизации между разными видами позднеплейстоценовых Homo (см. ссылки в новости Предки китайцев и японцев скрещивались с двумя разными популяциями денисовцев, «Элементы», 19.03.2018). Среди свежих новостей — обнаружение в геномах жителей Западной Африки значительной генетической примеси от неизвестной африканской популяции, которая обособилась, скорее всего, еще до разделения предков неандертальцев и сапиенсов (A. Durvasula, S. Sankararaman, 2020. Recovering signals of ghost archaic introgression in African populations). Ранее неоднократно сообщалось о загадочных архаичных примесях в геномах других африканских народов, в том числе пигмеев (P. Hsieh et al., 2016. Whole-genome sequence analyses of Western Central African Pygmy hunter-gatherers reveal a complex demographic history and identify candidate genes under positive natural selection).
Может быть, эпизоды межвидовой гибридизации и не были самым важным и интересным, что происходило в прошлом с людьми, но генетики как раз научились их выявлять с достаточной степенью надежности, а еще лет 15 назад это было невозможно — отсюда и повышенное внимание к этой теме в научной литературе.
Быстрый прогресс обеспечивается не только новыми отсеквенированными геномами, современными и древними, но и новыми методами их анализа. Исследование американских генетиков, о котором пойдет речь, основано как раз на применении нового метода к уже известным геномам.
Недавно Алан Роджерс (Alan R. Rogers), специалист по популяционной генетике и биоинформатике из Университета Юты, опубликовал описание пакета программ Legofit, разработанного им для реконструкции древней истории популяций по геномным данным (A. R. Rogers, 2019. Legofit: estimating population history from genetic data). В Legofit реализован новый подход к построению таких реконструкций, учитывающий накопленный опыт и свободный от некоторых погрешностей, характерных для других методов.
Главная идея состоит в том, что надежную информацию о древних событиях можно извлечь из частоты встречаемости различных комбинаций примитивных и продвинутых аллелей в разных нуклеотидных позициях («nucleotide site patterns») в геномах изучаемых популяций.
Допустим, у нас есть генетические данные по четырем популяциям (рис. 1). В данном случае это современные африканцы (обозначены буквой X), современные европейцы (Y), неандертальцы (N) и денисовцы (D). Для каждой нуклеотидной позиции (сайта) берется по одному случайно выбранному гаплоидному генотипу из каждой популяции. Внутрипопуляционная изменчивость при этом намеренно игнорируется, что позволяет абстрагироваться от недавних событий в истории популяций и сосредоточиться на древних (подробнее см. в упомянутой выше статье A. R. Rogers, 2019. Legofit: estimating population history from genetic data). Учитываются сайты, в которых у одних популяций оказался примитивный аллель, а у других — производный. Соответственно, для каждого сайта получается та или иная комбинация или рисунок (pattern) распределения примитивных и продвинутых аллелей. Например, может оказаться, что у первых трех популяций в данном сайте стоит такой же нуклеотид, как у шимпанзе (примитивный аллель), а у денисовцев — другой (производный аллель). Такая ситуация обозначается буквой d (рис. 1). Или, к примеру, может оказаться, что у первых трех популяций аллель производный и только у денисовцев — примитивный (комбинация xyn).
На рисунке 1 показано генеалогическое дерево изучаемых популяций (S — неизвестная «сверхархаичная» популяция, superarchaics; ее так назвали, потому что неандертальцев и денисовцев называют «архаичными людьми») с четырьмя эпизодами привноса чужих генов в результате гибридизации (стрелки с греческими буквами). Внутри ветвей дерева красными, синими и пунктирными линиями показана генеалогия одного конкретного нуклеотидного сайта. Если мутация произошла где-нибудь на синей ветке, то для данного сайта получится комбинация производных и примитивных аллелей xyn, а если на красной — d.
Ожидаемая частота встречаемости различных комбинаций зависит от параметров генеалогического дерева, а именно от порядка ветвления, длины ветвей, числа эпизодов гибридизации, их интенсивности (доли привнесенной ДНК), динамики численности рассматриваемых популяций. Программа Legofit подбирает эти параметры таким образом, чтобы ожидаемые частоты встречаемости разных комбинаций примитивных и производных аллелей были максимально похожими на реально наблюдаемые.
В статье, опубликованной 20 февраля в журнале Science Advances, Роджерс и его коллеги использовали этот метод для реконструкции популяционной истории сапиенсов, неандертальцев и денисовцев. Неандертальцы были представлены геномами двух индивидов: из Денисовой пещеры на Алтае и из пещеры Виндия в Хорватии. Эти два неандертальских генома отсеквенированы наиболее качественно. Они представляют довольно далекие друг от друга популяции неандертальцев. Для денисовцев есть только один качественно прочтенный ядерный геном, которым и была представлена денисовская популяция. Современную европейскую популяцию представляли геномы пяти французов и двух англичан, африканскую — геномы трех йоруба. В качестве внешней группы (для определения того, какие аллели являются примитивными) использовались геномы шимпанзе и гориллы.
Реальные (наблюдаемые) частоты встречаемости разных комбинаций примитивных и производных аллелей в рассматриваемых популяциях показаны на рис. 2.

Рис. 2. Частоты встречаемости разных комбинаций производных и примитивных аллелей у современных африканцев (x), современных европейцев (y), хорватского неандертальца (v), алтайского неандертальца (a) и денисовца (d). Рисунок из обсуждаемой статьи в Science Advances
Самое общее представление об искомом дереве можно получить, просто разглядывая рисунок. Если не учитывать «синглетоны» (комбинации, в которых производный аллель есть только у одной из пяти популяций), то чаще всего встречается комбинация va, роднящая алтайских и хорватских неандертальцев, и комбинация xy, отражающая родство современных европейских и африканских сапиенсов. Стало быть, это концевые веточки дерева, разделившиеся в последнюю очередь. На третьем месте — комбинация vad, говорящая о родстве неандертальцев и денисовцев (они ближе друг к другу, чем к сапиенсам). Значит, предковая популяция (ее можно обозначить как XYVAD) cначала разделилась на ветви XY (сапиенсы) и VAD (общие предки неандертальцев и денисовцев, которых авторы называют «неандерсовцами»). Затем ветвь VAD разделилась на VA и D (неандертальцев и денисовцев), VA — на V и A, XY — на X и Y. Это — давно известная топология родственных связей сапиенсов, неандертальцев и денисовцев.
Чтобы продвинуться дальше, уже требуется моделирование. Сначала (еще в своих прежних публикациях) авторы испытывали модели, вообще не учитывающие гибридизацию. Такие модели не в состоянии объяснить наблюдаемую картину (то есть воспроизвести частоты вариантов, показанные на рис. 2).
Добавление в модель одного эпизода гибридизации (привноса генов от неандертальцев к предкам европейцев — этот эпизод обозначен на рис. 1 буквой α) улучшило ситуацию, но не радикально. Тогда авторы добавили еще два выявленных ранее эпизода, β и γ. Эпизод β — это привнос генов от неизвестной архаичной популяции (S на рис. 1) в генофонд денисовцев, а эпизод γ — это гибридизация предков алтайских неандертальцев с какими-то древними сапиенсами (см.: У алтайских неандертальцев найдены гены архаичных сапиенсов, а у денисовцев — гены гейдельбергских людей, «Элементы», 25.02.2016).
Добавление эпизодов β и (или) γ сильно улучшает модель, но всё-таки не позволяет удовлетворительно объяснить наблюдаемую картину. Например, модель αβγ (с тремя эпизодами гибридизации) выдаёт близкие к реальности частоты комбинаций лишь в том случае, если «сверхархаики» отделились от предков сапиенсов, неандертальцев и денисовцев целых 7 млн лет назад. Это, конечно, ерунда: не могли же денисовцы в самом деле скрещиваться с шимпанзе.
Авторы предположили, что модели для совпадения с реальностью не хватает еще одного эпизода гибридизации. Вот только кого и с кем?
Общие предки денисовцев и неандертальцев — «неандерсовцы» — отделились от предков сапиенсов, возможно, около 750 тысяч лет назад. Позднее, где-то около 600 тысяч лет назад, предполагаемые предки неандертальцев появились в Европе, неся с собой ашельские орудия. В Азии, по идее, в это время должны были расселяться предки денисовцев. Те и другие селились не на безлюдных территориях. В Евразии уже давно жили представители рода Homo — потомки первой волны мигрантов, ранних Homo erectus, впервые покинувших африканскую прародину не позднее 1,8 млн лет назад (датировка людей из Дманиси). «Неандерсовцы» вполне могли поскрещиваться с евразийскими эректусами еще до того, как разделились на предков неандертальцев и денисовцев.
Предположение выглядело правдоподобным, и авторы начали экспериментировать с моделями, предполагающими еще один эпизод гибридизации — δ (рис. 1). Они сравнивали восемь моделей, включающих эпизоды β, γ и δ в разных комбинациях (эпизод α, в реальности которого нет сомнений, был включен во все модели). Сравнение проводили при помощи изощренных статистических методов, специально адаптированных для данной задачи, с учетом того, что чем больше в модели параметров, тем проще ее подогнать к наблюдаемым фактам, но только такая подгонка может оказаться неадекватной (так называемая проблема «переподгонки», см. также overfitting).
В результате с большим отрывом победила модель αβγδ, включающая все четыре эпизода гибридизации. На втором месте по правдоподобию — модель αβδ (игнорирующая приток генов от архаичных сапиенсов к алтайским неандертальцам). Все остальные варианты получили нулевую поддержку, то есть, по-видимому, вообще не имеют права на существование.
Таким образом, результаты уверенно подтвердили установленный ранее эпизод гибридизации β (приток генов от «сверхархаиков» к денисовцам) и чуть менее уверенно — эпизод γ (приток генов от архаичных сапиенсов к алтайским неандертальцам). Ну а самое главное, они свидетельствуют в пользу еще одного эпизода гибридизации δ, связанного с притоком генов от сверхархаиков к неандерсовцам. Это — новая гипотеза, такой эпизод ранее всерьез не обсуждался. Если он подтвердится дальнейшими исследованиями, это будет самый древний известный эпизод гибридизации между удаленными ветвями человеческого рода.
Дополнительные статистические тесты показали, что эпизод δ не может быть артефактом, порожденным ошибками секвенирования или соматическими мутациями.
На рисунке 3 показаны усредненные параметры генеалогического дерева, вычисленные програмой Legofit для двух наилучших моделей αβγδ и αβδ. При усреднении использовались «веса» этих моделей, отражающие их соответствие реальным данным (у первой модели вес 0,84, у второй 0,16). Проще говоря, эти параметры в основном соответствуют модели с четырьмя эпизодами гибридизации, αβγδ.

Рис. 3. Параметры популяционной истории сапиенсов, неандертальцев и денисовцев, оцененные при помощи программы Legofit. Верхний график показывает долю привнесенной ДНК для каждого из четырех эпизодов гибридизации (α — от неандертальцев к предкам современных европейцев, β — от сверхархаиков к денисовцам, γ — от древних сапиенсов к неандертальцам, δ — от сверхархаиков к общим предкам неандертальцев и денисовцев (неандерсовцам). Средний график показывает время важнейших событий (TXYNDS — отделение сверхархаиков от предков сапиенсов, неандертальцев и денисовцев, TXY — дивергенция предков современных африканцев и европейцев, TND — разделение неандертальцев и денисовцев, TN0 — конец раннего периода истории неандертальцев, когда их популяция была многочисленной, TA — время жизни алтайского неандертальца, TV — время жизни хорватского неандертальца, TD — время жизни денисовца (Denisova 3). Нижний график показывает оценки численности популяций (NS — сверхархаики, NXYND — предки сапиенсов, неандертальцев и денисовцев, NXY — предки европейцев и африканцев, NND — неандерсовцы, NN0 — ранние неандертальцы, NN1 — поздние неандертальцев). Рисунок из обсуждаемой статьи в Science Advances
У параметров, показанных на рис. 3, двоякий смысл. Если параметр ранее был оценен независимыми методами и новая оценка похожа на старую, то это прибавляет достоверности модели αβγδ, заодно подкрепляя и новую идею о скрещивании неандерсовцев со сверхархаиками. Так обстоит дело, например, с датировками костей денисовца и двух неандертальцев: они похожи на полученные ранее (см.: Уточнены датировки археологических находок в Денисовой пещере, «Элементы», 04.02.2019). Это — весомый аргумент в пользу достоверности модели αβγδ.
Если же новая оценка сильно отличается от прежних, то это указывает либо на несовершенство модели, либо на то, что прежние оценки нуждаются в уточнении. В эту категорию попадает неожиданно древняя датировка расхождения предков неандертальцев и денисовцев: 737 тысяч лет назад. Раньше давались оценки 200–300 тысяч лет назад (но это было давно), в последнее время чаще встречались оценки 400–500 тысяч лет назад. Но всё же не 700 с лишним! С одной стороны, это вроде бы слегка подрывает доверие к выводам авторов, с другой — более древняя датировка разделения неандертальцев и денисовцев лучше согласуется с древним возрастом людей из Симы де лос Уэсос, которые являются представителями неандертальской линии и должны были жить уже после разделения (см.: Семнадцать черепов из испанской пещеры проливают свет на происхождение неандертальцев, «Элементы», 24.06.2014).
Если же значение параметра ранее не имело обоснованных оценок, то оценка, показанная на рис. 3, представляет собой просто некий новый научный результат. К этой категории относится, например, время отделения сверхархаиков от предков сапиенсов, неандертальцев и денисовцев — 2,3 млн лет назад. Правда, нужно иметь в виду, что эта оценка (как и другие датировки, показанные на среднем графике на рис. 3) зависит от калибровки молекулярных часов, которая, в свою очередь, определяется темпом мутагенеза у наших предков, а на этот счет у специалистов пока нет консенсуса. Скорость мутагенеза (в расчете на нуклеотид в год) сильно зависит, например, от возраста мужского полового созревания, который не так-то просто определить для древних Homo. Авторы использовали относительно низкую оценку темпа мутагенеза (0,38·10−9 на нуклеотид в год), и это отчасти объясняет, почему некоторые датировки у них получились подозрительно древними. Если же взять более высокую (и не менее правдоподобную) оценку 0,45·10−9, то сверхархаики отделились от наших предков значительно позже — 1,9 млн лет назад.
Впрочем, обе эти датировки хорошо согласуются с предположением о том, что сверхархаики, с которыми скрещивались сначала неандерсовцы (эпизод δ), а потом денисовцы (эпизод β), были потомками самых первых выходцев из Африки — ранних евразийских эректусов, древнейшие представители которых нам известны по находкам из Дманиси (возраст 1,85–1,77 млн лет, см.: Пятый череп из Дманиси показал огромный размах индивидуальной изменчивости ранних Homo, «Элементы», 21.10.2013).
К относительно новым выводам (хотя отдельные намеки на них были и раньше) следует отнести очень низкую численность популяции неандерсовцев (NND на нижнем графике на рис. 3), высокую численность ранних неандертальцев по сравнению с поздними, а также очень высокую численность популяции сверхархаиков. Впрочем, к этим оценкам численности нужно относиться крайне осторожно. Достоверность их невысока, а главное, они могут на самом деле отражать не численность, а структуру популяции. Например, якобы огромная численность сверхархаиков может с тем же успехом означать, что евразийские эректусы рано подразделились на две ветви, одна из которых скрещивалась с неандерсовцами, а другая — с денисовцами. При этом обе ветви могли быть малочисленными. Главное, что они успели накопить много генетических различий. Ведь по сути дела речь идет о том, что две порции аллелей, доставшиеся от сверхархаиков другим людям (сначала неандерсовцам, потом денисовцам), были довольно разными. Это могло быть связано как с многочисленностью сверхархаиков, так и с тем, что они были представлены двумя рано разошедшимися популяциями.
Если выводы авторов верны, то складывается довольно красивая картинка. Для объяснения имеющихся генетических данных достаточно всего трёх волн выхода людей из Африки. Первыми вышли ранние эректусы (возможно, близкие к людям из Дманиси) с олдувайскими орудиями. Произошло это примерно 1,9–1,8 млн лет назад. Эректусы постепенно расселились по Евразии, возможно, разделившись на две большие ветви.
Тем временем в Африке неандерсовцы отделились от предков сапиенсов и вышли из Африки с ашельскими орудиями, может быть, около 0,8–0,6 млн лет назад. Они прошли через бутылочное горлышко численности, поскрещивались с местными эректусами (около 0,75 млн лет назад, если верить применяемой в работе замедленной версии молекулярных часов), а потом расселились по Евразии, разделившись на две ветви: западную и восточную, неандертальцев и денисовцев. В ходе своей экспансии они, по-видимому, сильно потеснили местных аборигенов — эректусов. Но всё же не довели их до полного вымирания, а оставили немного на развод: ведь денисовцы позднее опять скрещивались с эректусами.
История повторилась 60–50 тысяч лет назад, когда популяция сапиенсов вышла из Африки, прошла бутылочное горлышко численности, поскрещивалась с местными аборигенами — ближневосточными неандертальцами, а потом расселилась по Евразии, разделившись на две ветви (восточную и западную) и быстро вытеснив своих предшествеников, неандертальцев и денисовцев. При этом восточная ветвь успела поскрещиваться с денисовцами.
Если вывод о гибридизации неандерсовцев со сверхархаиками верен, то это не только самый древний документированный эпизод отдаленной гибридизации в роде Homo, но и эпизод, участники которого прожили в изоляции друг от друга исключительно долго — более миллиона лет. Ну а когда денисовцы повторно скрещивались со сверхархаиками, время раздельной жизни гибридизующихся популяций уже составляло порядка полутора миллионов лет. Это не рекордное, но весьма солидное время раздельной жизни для межвидовой гибридизации у млекопитающих (см. рис. 2 в новости Плацентарность не способствует мужской красоте, «Элементы», 16.09.2014).
Источник: elementy.ru





