













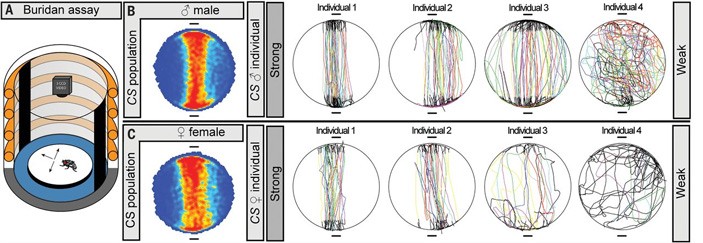


Рис. 1. Схема эксперимента по выявлению индивидуальных поведенческих особенностей у дрозофил (Buridan assay — «буридановский» тест). Муху с обрезанными крыльями сажают на круглую, ярко освещенную, окруженную водой арену. В такой ситуации мухе очень хочется оказаться на одной из двух вертикальных черных полос, расположенных на светящейся стенке. Однако ров с водой не дает мухе до них добраться. Убедившись в недосягаемости одной из полос, муха разворачивается и идет к другой, и так до бесконечности (на самом деле каждый тест длился 900 секунд, но известно, что муха в этой ситуации может ходить туда-сюда тысячи раз, вплоть до полного истощения). Индивидуальные поведенческие особенности проявляются в том, что один особи ходят почти по прямой (сильная ориентация на объект, Strong), а другие то и дело отклоняются от прямого курса (слабая ориентация, Weak). Показаны усредненные траектории для самцов и самок мух из линии Canton S (CS population), а также индивидуальные маршруты четырех самцов и четырех самок. Рисунок из обсуждаемой статьи в Science
Индивидуальные поведенческие особенности животных теоретически могут объясняться не только генами и средой (воспитанием, условиями развития), но и случайной изменчивостью в строении мозга, которая есть даже у генетически идентичных особей, развивавшихся в одинаковых условиях. Формирование поведенческой индивидуальности на основе случайных флуктуаций в развитии мозга удалось в деталях изучить на дрозофилах — модельных животных, для которых разработан богатейший арсенал генетических и нейробиологических исследовательских инструментов. Оказалось, что индивидуальная изменчивость мух по точности ориентации на визуальные стимулы тесно связана со случайной изменчивостью по степени асимметрии зрительных областей мозга: чем сильнее асимметрия, тем точнее ориентация. Асимметрия формируется на определенной стадии развития мозга, когда нейроны «выбирают», куда им посылать свои отростки, причем в этом выборе присутствует элемент случайности.
Все животные, включая людей, ведут себя по-разному. Даже если у двух животных одинаковые гены (как у однояйцевых близнецов) и развитие шло в одинаковых условиях, они всё равно будут хоть немного, но отличаться друг от друга, в том числе и по поведению.
Все привыкли трактовать поведенческую индивидуальность в рамках дихотомии «гены или среда» (Nature versus nurture), хотя эта модель принципиально неполна. Кроме генов и среды есть еще один важный источник изменчивости, о котором вроде бы все знают, но часто забывают. Это случайные флуктуации, неизменно присутствующие на всех уровнях организации живых систем и проявляющиеся в любых биологических процессах, в том числе в онтогенезе (developmental noise; см. ссылки в новости Случайная изменчивость генной экспрессии у Arabidopsis thaliana подчиняется строгим закономерностям, «Элементы», 29.01.2019).
В строении мозга точно есть случайная изменчивость, и она должна, по идее, как-то влиять на поведение. Однако прямые экспериментальные подтверждения такого влияния получить не так-то просто. В новом исследовании, результаты которого опубликованы в журнале Science, группе биологов из Франции, Германии и Бельгии впервые удалось продемонстрировать четкую связь между случайными флуктуациями в анатомии мозга и индивидуальными поведенческими особенностями. Работа выполнена на мушках-дрозофилах, однако полученные выводы наверняка в какой-то степени приложимы и к другим животным.
Основное внимание в работе уделено так называемым «нейронам дорсального кластера» (dorsal cluster neurons, DCNs), которые участвуют в обработке зрительной информации. Нейроны DCN располагаются двумя группами, слева и справа, на дорсальной (спинной) стороне мозга. Они посылают сигналы в две зрительные области противоположного полушария, которые называются медулла (medulla) и лобула (lobula) (рис. 2).

Рис. 2. Нейроны DCN и зрительные зоны мушиного мозга. Тела нейронов DCN (Cell bodies) находятся на спинной стороне мозга слева и справа. От каждого нейрона отходит ипсилатеральный (остающийся в том же полушарии) дендрит и контралатеральный (идущий в противоположное полушарие) аксон. Эти аксоны образуют комиссуру — перемычку между двумя полушариями. Каждый аксон иннервирует либо лобулу (Lobula axons and dendrites), либо медуллу (Medulla axons). Нейроны M-DCN, аксоны которых иннервируют медуллу, покрашены зеленым. Красным покрашены все нейроны DCN. Длина масштабной линейки 20 мкм. Изображение из обсуждаемой статьи в Science
В процессе развития каждый аксон DCN «выбирает», в какую из двух областей ему направить свой аксон, причем в этом выборе присутствует элемент случайности. Соответственно, нейроны DCN подразделяются на две популяции: те, что иннервируют медуллу (M-DCN) и те, что иннервируют лобулу (L-DCN). Механизм выбора нейронами DCN одной из двух целевых областей хорошо изучен, и утверждение о том, что в выборе присутствует элемент случайности, имеет под собой солидный научный базис (M. Langen et al., 2013. Mutual inhibition among postmitotic neurons regulates robustness of brain wiring in Drosophila). Таким образом, зрительные зоны мушиного мозга — это пример неплохо изученной нейронной сети, в «прошивке» которой имеется ярко выраженная случайная изменчивость, причем природа этой изменчивости (почему и на каком этапе онтогенеза возникает «шум») более или менее понятна. Авторы решили выяснить, влияет ли эта стохастическая изменчивость на индивидуальные особенности поведения мух.
Поведение оценивалось при помощи «буридановского» теста, который традиционно используется для изучения визуальной ориентации мух (рис. 1, A). Предварительные опыты показали, что и для самцов, и для самок дрозофил из стандартной лабораторной линии Canton S характерна сильная изменчивость по склонности отклоняться от прямого маршрута, соединяющего два привлекательных визуальных стимула. Одни особи ходят туда-сюда строго по прямой, другие немного отклоняются в стороны, третьи блуждают почти случайно. Степень отклонения от прямого маршрута (absolute stripe deviation, aSD) можно измерить и в дальнейшем анализировать изменчивость по этому признаку статистическими методами.
Для начала авторы оценили изменчивость по aSD у нескольких инбредных, генетически однородных линий дрозофил. Оказалось, что существуют линии мух с повышенной и пониженной (по сравнению с Canton S) средней точностью ориентации. Однако индивидуальная изменчивость по этому признаку всегда остается высокой — даже в линиях с крайне низким генетическим разнообразием. Более того, эксперименты по скрещиванию мух с самыми высокими и самыми низкими значениями aSD показали, что у потомства от таких скрещиваний изменчивость по aSD ничуть не уменьшается. Следовательно, эта изменчивость в основном не наследственная.
Дополнительные опыты показали, что одна и та же муха, если тестировать ее многократно в течение жизни, демонстрирует одинаковое поведение. Ученые даже пробовали морить мух голодом, а потом снова их откармливать: это меняет среднее значение признака aSD в популяции (голодные мухи ходят в среднем прямее, а отъевшиеся после голодовки — «кривее», чем контрольные). Однако индивидуальные черты каждой мухи при этом сохраняются. Те особи, которые ходят прямее других в обычном состоянии, сохраняют эту особенность и в радости, и в горе.
Таким образом, индивидуальные различия по aSD мало зависят от текущего физиологического состояния или настроения мухи. По-видимому, это устойчивая индивидуальная характеристика, врожденная, хоть и не наследственная. Такие поведенческие черты теоретически как раз и могут возникать из-за случайных флуктуаций при формировании межнейронных связей в развивающемся мозге. Поэтому авторы решили проверить, не связаны ли обнаруженные поведенческие различия с индивидуальной изменчивостью по иннервации лобулы и медуллы нейронами DCN. Эта гипотеза была взята не с потолка: ранее некоторые специалисты на основе поведенческих экспериментов (в том числе с мутантными мухами, лишенными тех или иных групп нейронов в зрительных областях мозга) уже высказывали предположение, что визуальная ориентация у дрозофил должна быть как-то связана, во-первых, с бинокулярным зрением и связью между полушариями (а аксоны нейронов DCN как раз образуют комиссуру между полушариями), во-вторых — с асимметрией правых и левых зрительных областей (а поскольку в развитии DCN присутствует элемент случайности, это может приводить в том числе и к асимметрии полушарий).
Нейробиология дрозофилы — наука передовая, располагающая внушительным арсеналом современных исследовательских инструментов. Можно окрашивать разные группы нейронов, в том числе M-DCN и L-DCN, в разные цвета методами иммуногистохимии (рис. 2), можно делать детальные реконструкции отделов мозга, прослеживать путь отдельных аксонов, избирательно подавлять активность тех или иных групп нейронов, аккуратно вмешиваться в развитие мозга (например, блокируя синаптическую передачу при помощи термочувствительного белка UAS-shibirets и нагревания куколки до 30°C в определенный момент ее развития), можно при помощи двухфотонного микроскопа разглядывать детали строения мозга у куколки, не убивая ее, что позволяет изучать возрастные изменения структуры мозга у одной и той же особи, и так далее.
Исследователи в полной мере воспользовались этим арсеналом. Для начала они изучили зрительные области мозга у 103 особей, подсчитав все нейроны DCN и проследив путь аксонов. Оказалось, что в каждом полушарии есть по 22–68 нейронов DCN, в том числе 11–55 L-DCN и 6–23 M-DCN. Кроме того, обнаружилась сильная индивидуальная изменчивость по асимметрии иннервации левой и правой медуллы нейронами M-DCN. Это значит, что у одних мух число аксонов M-DCN, приходящих в левую и правую медуллу, одинаковое, а у других разное, причем иногда — очень разное (рис. 3, B).
Авторы убедились, что аксоны M-DCN приходят как раз в ту часть медуллы, которая получает визуальную информацию от переднего поля бинокулярного зрения (а значит, может иметь отношение к визуальной ориентации). Подавление активности M-DCN (или сразу всех нейронов DCN) приводит к резкому увеличению aSD, то есть мухи фактически перестают ориентироваться на черные полоски и начинают блуждать по арене случайным образом. Это опять-таки согласуется с предположением о связи M-DCN с визуальной ориентацией. Кроме того, авторы разглядывали мозги у живых куколок, а потом у вышедших из этих куколок мух. Это позволило убедиться, что иннервация медуллы нейронами DCN, сложившаяся на стадии куколки, в дальнейшем уже не меняется.
Но самый важный результат был получен, когда ученые сопоставили строение зрительных зон у упомянутых 103 мух с их поведением (которое, конечно, было тщательно записано перед тем, как отправить муху «под нож» для изучения мозга). Оказалось, что между aSD и асимметрией иннервации медуллы есть четкая корреляция: чем симметричнее иннервация, тем выше aSD. Иными словами, мухи с сильной асимметрией мозга в «буридановском» тесте ходят по прямой, а мухи с симметричным мозгом склонны к извилистым маршрутам (рис. 3).

Рис. 3. Асимметрия иннервации медуллы нейронами M-DCN положительно коррелирует с четкостью визуальной ориентации в «буридановском» тесте. А — общая схема эксперимента. Сначала записывают поведение мухи в «буридановском» тесте (Behaviour), затем с помощью иммуногистохимического окрашивания, конфокальной микроскопии и специального программного обеспечения получают реконструкцию зрительных отделов мозга (Anatomical recording, Neural reconstructions) и, наконец, ищут связь между поведением и строением мозга (Comparison Behaviour / Neural reconstructions). B — иннервация медуллы нейронами M-DCN и поведение двух произвольно выбранных мух. У «верхней» мухи иннервация резко асимметрична: в левую медуллу приходят аксоны от восемнадцати M-DCN, а в правую — только от двенадцати. Эта муха неплохо ориентировалась на визуальные стимулы в «буридановском» тесте. У «нижней» мухи иннервация симметричная (по 13 аксонов с каждой стороны), а визуальная ориентация выражена намного слабее. C — отрицательная корреляция между асимметрией медуллы (по вертикальной оси) и aSD, то есть отклонением от прямого маршрута (по горизонтальной оси). Чем сильнее асимметрия, тем прямее ходит муха. Синим и красным крестиками показаны две мухи с рисунка B. Изображение из обсуждаемой статьи в Science
Авторы провели ряд дополнительных экспериментов, чтобы убедиться, что случайная изменчивость иннервации медуллы нейронами M-DCN действительно является причиной индивидуальных поведенческих особенностей. Все проверки дали положительный результат: инактивация M-DCN приводит к исчезновению корреляции между асимметрией мозга и поведением; для инбредных линий с высоким средним aSD характерна пониженная (тоже в среднем) асимметрия мозга и наоборот. Авторам даже удалось вмешаться в развитие нейронов DCN на той ключевой стадии, когда они отращивают аксоны, обмениваясь друг с другом сигналами, что и приводит в конечном счете к асимметрии полушарий. Эти вмешательства привели к снижению среднего уровня асимметрии, и мухи в итоге хуже ориентировались на визуальные стимулы. Были проведены также генно-инженерные манипуляции, позволившие при помощи белка Kir2.1 инактивировать нейроны DCN только слева или только справа, что в итоге дало мух с искусственно усиленной асимметрией полушарий. Такие мухи в «буридановском» тесте ходили ровнее контрольных.
Таким образом, исследование показало, как случайные флуктуации в развитии мозга могут приводить к сохраняющимся на всю жизнь индивидуальным особенностям поведения. Теоретически такой механизм предполагался давно, но в деталях проследить его работу удалось впервые. Хотя это сделано пока только на дрозофилах, авторы уверены, что стохастика в развитии мозга должна вносить важный вклад в формирование поведенческой индивидуальности и у других животных, включая людей. С эволюционной точки зрения это можно рассматривать как вариант «биологического хеджирования ставок» (biological bet hedging): в переменчивой, плохо предсказуемой среде случайная изменчивость поведения может повышать вероятность того, что хотя бы кто-то из обладателей данного генотипа в критической ситуации поведет себя «правильно» и благодаря этому выживет и оставит потомство. В связи с этим вспоминаются антропологические данные об удивительно большом размахе индивидуальной изменчивости по объему мозга у представителей рода Homo (см.: Пятый череп из Дманиси показал огромный размах индивидуальной изменчивости ранних Homo, «Элементы», 21.10.2013): как знать, может быть и эта изменчивость была во многом случайной и служила для повышения интеллектуального и поведенческого разнообразия у наших предков?
Источник: elementy.ru





