













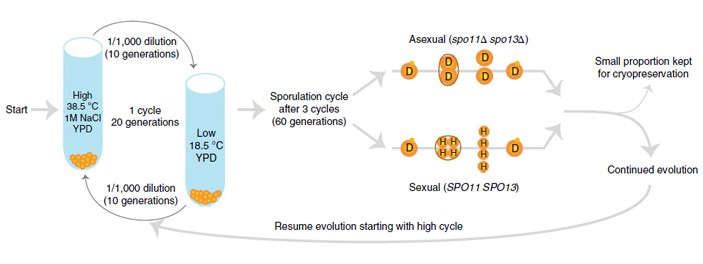


Рис. 1. Схема эксперимента. Использовались два штамма диплоидных дрожжей, одинаковых во всём, кроме способности к половому размножению: «половой» штамм (Sexual (SPO11 SPO13)) и «бесполый» (Asexual (spo11Δ spo13Δ)). У бесполых дрожжей в условиях, провоцирующих споруляцию, вместо четырех гаплоидных спор образуются две диплоидные клетки, то есть вместо мейоза происходит обычный митоз. Из каждого штамма сделали по 6 подопытных популяций. Их содержали попеременно то при высокой температуре (38,5°C) в соленой среде, то при низкой температуре (18,5°C) в среде без добавления соли. Условия менялись каждые 10 поколений, полный цикл занимал 20 поколений. После каждых трех таких циклов, то есть каждые 60 поколений, дрожжи помещались в специальную среду, которая провоцирует споруляцию (Sporulation cycle after 3 cycles (60 generations)). «Половые» дрожжи при этом претерпевали мейоз, а получившиеся гаплоидные разнополые клетки (H) потом спаривались, сливаясь снова в диплоидные (D). У бесполых дрожжей споруляция сразу приводила к формированию диплоидных клеток. В обоих случаях получившиеся диплоидные клетки затем приступали к следующему циклу адаптации (Continued evolution). Эксперимент продолжался 24 полных цикла, то есть примерно 1440 поколений. Рисунок из обсуждаемой статьи в Nature Ecology & Evolution
При бесполом размножении отбор может работать только с целыми геномами, но не с отдельными генами. Из-за этого многие слабовредные мутации распространяются вместе с полезными («генетический автостоп»), а полезные мутации, возникшие в разных геномах, не могут объединиться и конкурируют друг с другом («клональная интерференция»). Половой процесс сводит к минимуму генетический автостоп и клональную интерференцию, тем самым помогая организмам быстрее адаптироваться к меняющимся условиям среды. Ранее в эволюционном эксперименте на гаплоидных дрожжах эти теоретические представления блестяще подтвердились. Новое исследование, проведенное по сходной методике, показало, что у диплоидных дрожжей всё немного сложнее. Бесполые диплоидные популяции адаптировались не хуже популяций, способных к половому процессу. Как выяснилось, успешная адаптация бесполых диплоидов отчасти связана с распространением мутаций, полезных только в гетерозиготном состоянии, но бесполезных или вредных в гомозиготном (это называют «сверхдоминированием»). Половой процесс лишает эволюционирующую популяцию этой опции, потому что потомство от скрещивания двух гетерозигот наполовину состоит из гомозигот. Возможность адаптироваться за счет «сверхдоминирующих» мутаций отчасти компенсирует бесполым диплоидам ущерб, причиняемый генетическим автостопом и клональной интерференцией.
В 2016 году в журнале Nature была опубликована статья, которая, казалось, расставила все точки над «i» в вопросе о том, как влияет половой процесс на эволюцию геномов и почему он помогает организмам быстрее адаптироваться к условиям среды. Об этом впечатляющем исследовании рассказано в новости Половое размножение помогает отбору отделять полезные мутации от вредных («Элементы», 01.03.2016). Эволюционный эксперимент проводился на 18 популяциях гаплоидных дрожжей, из которых 12 размножались только клонально (почкованием), а остальные шесть периодически (раз в 90 поколений) перекомбинировали свои гены в ходе полового процесса.
За 1000 поколений половые популяции приспособились к условиям эксперимента значительно лучше бесполых. Анализ геномов показал, что в половых популяциях закрепились только полезные мутации, тогда как в бесполых популяциях накопилось много «мусора», то есть нейтральных и слабовредных мутаций. Таким образом, подтвердилась идея о том, что одной из причин менее эффективной адаптации клональных популяций является генетический автостоп (Genetic hitchhiking) — распространение неудачных генетических вариантов вместе с полезными мутациями, поддержанными отбором. Без полового процесса полезную мутацию невозможно отделить от всего остального, плохого и хорошего, что есть в том геноме, где эта полезная мутация возникла.
Анализ изменения частоты встречаемости новых аллелей, возникших в ходе эксперимента, подтвердил и второе важное теоретическое предсказание — о том, что в бесполых популяциях происходит «клональная интерференция» (Clonal interference), то есть безжалостная конкуренция между потомками организмов с разными полезными мутациями. В отличие от половых популяций, где такие мутации быстро объединяются в одном геноме, в бесполых популяциях они не могут объединиться и вместо этого конкурируют, вытесняя друг друга.
Однако эти замечательные результаты пока были получены только на гаплоидных популяциях дрожжей. В природе же большинство эукариот предпочитает жить в диплоидном состоянии, что усложняет ситуацию. Вместе с диплоидностью появляются, например, такие явления, как доминантность и рецессивность, что не может не сказаться на молекулярной эволюции.
Пекарские дрожжи Saccharomyces cerevisiae — удобнейший объект для оценки влияния полового размножения на эволюцию гаплоидных и диплоидных популяций. Дрожжи могут размножаться почкованием как в гаплоидной, так и в диплоидной фазе своего жизненного цикла (см. рис. 1 в вышеупомянутой новости). Половой процесс у дрожжей состоит из двух этапов — слияние двух гаплоидных клеток, относящихся к разным «полам» а и α, и мейоз диплоидной клетки, приводящий к формированию четырех гаплоидных спор, — причем оба этапа можно аккуратно контролировать в лаборатории. Говоря упрощенно, чтобы гаплоидные клетки не спаривались, нужно просто содержать разнополые клетки отдельно друг от друга, а споруляцию можно стимулировать, помещая диплоидные клетки в специальную среду. Чтобы было удобно отделять гаплоидные клетки от диплоидных, а пол а от пола α, в экспериментах используются специально выведенные штаммы дрожжей, которые в зависимости от своей плоидности и пола могут или не могут расти на разных питательных средах, например, содержащих или не содержащих урацил и лейцин (подробнее об этом рассказано в упомянутой выше новости).
Итак, напрашивается идея проверить на диплоидных дрожжах выводы о роли полового размножения, полученные ранее на гаплоидных дрожжах. Именно это и попытались сделать Майкл Макдональд (Michael J. McDonald) из Университета Монаша (Австралия), его аспирантка Лора Вудс (Laura Woods) и их коллеги из Академии Синика (Academia Sinica, Тайвань). Статья с результатами их опытов опубликована 10 февраля в журнале Nature Ecology & Evolution. В публикации 2016 года Макдональд — первый автор. Тогда он был постдоком в Гарварде. Сейчас он завлаб в Монаше, и в новой статье его фамилия стоит на почетном последнем месте. Все остальные авторы в двух статьях разные.
Общая схема эволюционного эксперимента показана на рис. 1. Важно иметь в виду, что отличия от эксперимента 2016 года не сводятся к плоидности дрожжей, и поэтому результаты двух экспериментов, к сожалению, не очень просто сравнивать.
Важное отличие состоит в том, что эксперимент на гаплоидных дрожжах проводился в постоянных и относительно благоприятных условиях, а на диплоидных — в переменных и неблагоприятных. Дрожжи культивировались то при высокой температуре (38,5°C) в соленой среде (молярная концентрация 1M — около 6% NaCl), то при низкой (18,5°C) в среде без добавления соли. Оба режима для дрожжей являются неблагоприятными: оптимальная температура их культивирования в лаборатории 30°C. Поэтому, сравнивая результаты двух экспериментов, нужно помнить, что априори не известно, с чем связаны обнаруженные различия — с плоидностью или с разными условиями, к которым приходилось приспосабливаться дрожжам. Кроме того, циклы полового размножения в новом эксперименте осуществлялись каждые 60 поколений, а в 2016 году — каждые 90 поколений. Но хотя бы численности популяций в обоих экспериментах были примерно одинаковыми (105 в моменты «бутылочных горлышек», когда клетки переселялись в новую среду).
По окончании эксперимента, который продолжался примерно 1440 поколений (в 2016 году — 1000 поколений), ученые сравнили приспособленность (скорость роста) подопытных популяций и исходных (предковых) дрожжей. Сравнение проводилось по отдельности в «горячих» условиях, в «холодных» и в переменных.
Результаты показаны на рис. 2 (a–c). К «горячим» условиям хорошо приспособились все 12 популяций (рис. 2а). Напротив, приспособленность к «холодным» условиям в большинстве популяций осталась на исходном уровне, а у одной половой популяции даже ухудшилась (рис. 2b). Возможно, это связано с тем, что в «горячих» условиях дрожжи исходно чувствовали себя хуже, чем в «холодных». Иными словами, их изначальная приспособленность к «горячим» условиям была ниже. Между тем в нескольких эволюционных экспериментах было показано, что на ранних этапах адаптации приспособленность растет тем быстрее, чем ниже ее исходный уровень (см. Предсказуемый рост приспособленности достигается непредсказуемыми путями, «Элементы», 30.06.2014).

Рис. 2. Основные результаты. a–c — приспособленность 12 подопытных популяций по окончании эволюционного эксперимента. a1–a6 — бесполые, s1–s6 — половые популяции. a — приспособленность к «горячим» условиям, b — к «холодным» условиям, c — к полному экспериментальному циклу, когда надо прожить 10 поколений в горячих условиях и еще 10 — в холодных. В случаях «a» и «b» приспособленность подопытных популяций сравнивали с исходным (предковым) штаммом по результатам конкурентных тестов. В случае «с» исходный штамм совсем не смог конкурировать со своими проэволюционировавшими потомками, поэтому за точку отсчета взяли одну из наименее приспособленных бесполых линий (а1). По вертикальной оси показан коэффициент отбора (Selection coefficient). Если он равен нулю, это значит, что подопытная популяция размножается в данных условиях с той же скоростью, что и контрольная. Положительные значения указывают на рост приспособленности, отрицательные — на ее снижение. Ни в одной из трех сред (a, b, c) между приспособленностью половых и бесполых популяций не обнаружилось значимых различий (NS — not significant). d — количество обнаруженных мутаций в половых (Sexual) и бесполых (Asexual) популяциях. All — все зарегистрированные мутации, Het — мутации с частотой встречаемости от 0,4 до 0,9, которые, скорее всего, присутствуют в популяции в основном в гетерозиготном состоянии, Hom — мутации с частотой более 0,9, присутствующие в популяции в основном в гомозиготном состоянии. Разными цветами показаны несинонимичные (Non-synonymous), синонимичные (Synonymous) и межгенные мутации (Intergenic). Рисунок из обсуждаемой статьи в Nature Ecology & Evolution
Приспособленность к переменным условиям эксперимента сильно выросла у всех подопытных популяций по сравнению с предковым штаммом — настолько, что тот вообще не смог конкурировать со своими потомками в проверочных тестах. Поэтому за «точку отсчета» пришлось взять одну из наименее приспособленных подопытных популяций, а1 (рис. 2с).
Самое интересное, что ни в одной из трех ситуаций не удалось выявить значимых различий между приспособленностью половых и бесполых популяций. Иными словами, хваленые преимущества полового размножения, о которых так долго говорили эволюционисты-теоретики, в этом эксперименте (в отличие от множества других) почему-то не проявились! Бесполые популяции за 1440 поколений приспособились к условиям эксперимента не хуже, чем половые.
В чем тут дело? Чтобы разобраться, исследователи обратились к анализу геномов. Они отсеквенировали пробы ДНК каждой проэволюционировавшей популяции, чтобы обнаружить новые мутации, возникшие в ходе эксперимента, и оценить частоту их встречаемости. При этом ДНК брали не из отдельных клеток или клонов, а из всей популяции в целом. Недостатком этого подхода является то, что он не позволяет напрямую оценить количество гомо- и гетерозигот. Например, если частота встречаемости мутации равна 0,5, то это может означать, что все клетки в популяции являются гетерозиготами, либо что в ней половина клеток гомозиготны по наличию мутации, а остальные — по ее отсутствию, со всеми возможными промежуточными вариантами (включая и строгое соблюдение закона Харди — Вайнберга). Конечно, в бесполой популяции частота 0,5 скорее всего говорит о всеобщей гетерозиготности, а в половой логично предположить соблюдение закона Харди — Вайнберга, но это, строго говоря, еще надо доказывать, чего авторы сделать не могли.
Результаты показаны на рис. 2d. Они во многом похожи на результаты исследования 2016 года (рис. 2 в новости Половое размножение помогает отбору отделять полезные мутации от вредных). Ясно, что у диплоидных дрожжей, как и у гаплоидных, половое размножение позволило отбору по отдельности работать с полезными и вредными мутациями. Поэтому в итоге в половых популяциях закрепилось (или достигло значительной частоты) меньшее число мутаций, однако все они, по-видимому, были полезными (дальнейшие эксперименты это подтвердили). В бесполых популяциях из-за генетического автостопа вместе с полезными мутациями распространялся и весь их «генетический фон». Это привело к распространению нейтральных, а возможно, и вредных мутаций. Это видно, в частности, по тому, что в бесполых популяциях распространилось намного больше синонимичных и межгенных (то есть, скорее всего, нейтральных) мутаций, чем в половых популяциях.
Неожиданным результатом является большое число мутаций, достигших частоты ≥0,9 в бесполых популяциях. Такая высокая частота означает, что большинство клеток в популяции являются гомозиготными по данной мутации. Но как может мутация стать гомозиготной без полового процесса? Ведь вероятность того, что одна и та же мутация случайно возникнет в обеих гомологичных хромосомах, пренебрежимо мала! Авторы поясняют, что это, скорее всего, результат митотического кроссинговера (Mitotic recombination), который у диплоидных дрожжей нередко приводит к потере гетерозиготности даже без полового процесса.
Характерным признаком адаптивной эволюции в экспериментах являются параллелизмы, то есть независимое распространение одинаковых мутаций в разных популяциях, адаптирующихся к одинаковым условиям (см. Параллельная эволюция изучена в эксперименте на бактериях, «Элементы», 01.02.2012). Судя по этому критерию, так же как и по количеству и качеству распространившихся мутаций, адаптивная эволюция в половых популяциях работала эффективнее, чем в бесполых. В половых популяциях обнаружилось целых 13 генов, мутации в которых распространились более чем в одной популяции, в том числе три гена, изменившиеся сразу в трех популяциях, и один ген, изменившийся в пяти популяциях. В бесполых популяциях нашлось лишь четыре гена, мутации в которых распространились в двух популяциях. Такой уровень эволюционного параллелизма статистически неотличим от случайного шума. Интересно, что мутации в четырех генах (SKY1, HRD3, LEU3, FKS1) были замечены только по одному разу в бесполых популяциях, а в половых — по два раза или более. Это значит, что мутации в этих генах, скорее всего, были полезны как половым, так и бесполым популяциям, но только в бесполых популяциях у отбора было меньше шансов их поддержать и довести до заметной частоты (из-за сцепленности с вредными мутациями и клональной интерференции).
Итак, судя по таким важным признакам, как характер распространившихся мутаций и наличие параллелизмов, половые популяции адаптировались эффективнее, чем бесполые. Почему же тогда их итоговая приспособленность оказалась примерно такой же?
По-видимому, дело тут в дополнительном эволюционном «бонусе», который получают диплоидные организмы, лишенные полового процесса. Бонус состоит в том, что они могут адаптироваться за счет мутаций, полезных только в гетерозиготном состоянии (см. Сверхдоминирование). В половых популяциях таким мутациям гораздо труднее достичь высокой частоты, потому что при скрещивании двух гетерозигот половина потомков оказываются гомозиготами и, следовательно, имеют пониженную приспособленность.
Косвенные указания на важную роль сверхдоминирования в эволюции бесполых диплоидов дал анализ изменений частот мутаций во времени, который авторы провели для трех половых и трех бесполых популяций (рис. 3).

Рис 3. Изменение частоты встречаемости мутаций в трех бесполых (a–c) и трех половых популяциях (d–f). По горизонтальной оси — поколения, по вертикальной — частота мутаций. Буквами обозначены гены, мутации в которых распространились более чем в одной популяции. Рисунок из обсуждаемой статьи в Nature Ecology & Evolution
Рисунок показывает, во-первых, что в бесполых популяциях частоты многих мутаций растут и снижаются синхронно. Синхронный рост указывает на генетический автостоп, а синхронные снижения — на потерю ценных мутаций в результате клональной интерференции. В половых популяциях мутации ведут себя независимо, у каждой — своя индивидуальная судьба.
Во-вторых, в бесполых популяциях у многих мутаций рост частоты встречаемости останавливается возле отметки 0,5. Это почти наверняка означает, что все или почти все клетки в популяции стали гетерозиготными по этой мутации. В половых популяциях ничего похожего не наблюдается.
Авторы изучили влияние нескольких мутаций на приспособленность дрожжей, искусственно внедряя эти мутации в геном предкового штамма. Из трех проверенных мутаций, распространившихся у бесполых дрожжей, две оказались нейтральными или вредными как в «горячих», так и в «холодных» условиях. Очевидно, эти ненужные мутации могли распространиться только благодаря генетическому автостопу. Третья мутация, выводящая из строя ген CDC39, оказалась полезной в «холодных» условиях, но только в гетерозиготном состоянии. В гомозиготном состоянии она летальна. Это яркий пример сверхдоминирования. Неудивительно, что в половой популяции s6, где эта мутация тоже появилась (рис. 3f), она не смогла достичь высокой частоты. Те же мутации, которые достигли высокой частоты в популяции s6 (URA2-2 и STP4), оказались полезными как в гетеро-, так и в гомозиготном состоянии.
Таким образом, исследование показало, что у диплоидных дрожжей, как и у гаплоидных, половой процесс помогает отбору отделять полезные мутации от вредных и надежно защищает популяцию от вредных последствий генетического автостопа и клональной интерференции. Но это не обязательно приводит к тому, что половые популяции адаптируются быстрее и лучше, чем бесполые, потому что диплоидность дает бесполым популяциям дополнительный бонус — возможность адаптироваться за счет мутаций, полезных только в гетерозиготном состоянии. В долгосрочной перспективе у бесполых диплоидов два аллельных варианта гена могут даже приобрести разные функции (см. Утрата полового размножения способствует появлению новых генов, «Элементы», 15.10.2007), хотя заметить такие процессы в лабораторном эксперименте мало шансов.
Исследование представляет собой лишь первую попытку детального сравнения молекулярной эволюции в половых и бесполых диплоидных популяциях, так что не стоит делать из него далеко идущих выводов. В методике эксперимента есть уязвимые места, да и в результатах кое-что настораживает. Например, среди 13 генов, мутации в которых распространились более чем в одной половой популяции, есть два гена, URA2 и LEU3, функции которых связаны с синтезом урацила и лейцина. Это выглядит подозрительно, потому что во время циклов полового размножения для отбора разнополых гаплоидных клеток использовались питательные среды, не содержащие урацила и лейцина. Этой процедуре подвергались только половые популяции. Поэтому можно предположить, что направленность и сила отбора, действовавшего на половые и бесполые популяции в ходе эксперимента, были не совсем одинаковыми. Может быть, половые популяции адаптировались на самом деле не к двум, а к трем разным наборам неблагоприятных условий: к холоду, жаре и вдобавок к периодическому дефициту лейцина и урацила. Это тоже могло помешать им обогнать бесполых конкурентов по приспособленности к жаре и холоду.
И всё же нельзя не порадоваться быстрому развитию исследовательских методов, позволяющих эволюционным биологам ставить и решать такие задачи, к которым еще лет 10 назад было невозможно подступиться.
Источник: elementy.ru





