













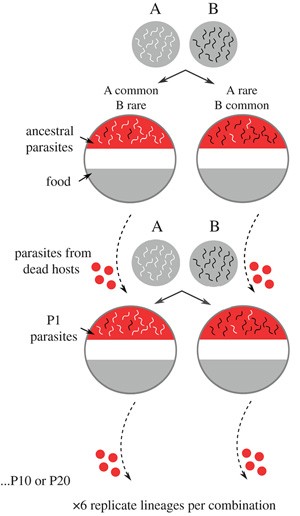


Рис. 1. Дизайн эксперимента. Один штамм бактерии Serratia marcescens использовали для создания 36 независимых линий. Эти линии прошли отбор на увеличение вирулентности (смертности хозяев) в гетерогенных популяциях нематод, в которых один генотип хозяина был частым, а другой — редким. В каждом раунде отбора не эволюционирующие хозяева добавлялись к паразитам, а через 24 часа бактерии извлекались из умерших хозяев. Эти паразиты (которые относительно быстро убили своих хозяев) переходили в следующий раунд отбора. Четыре генотипа нематод были объединены в пары в различных комбинациях и частотах, чтобы получилось шесть вариантов гетерогенных популяций хозяина. Для каждого сочетания генотипов использовали шесть повторностей. Рисунок из обсуждаемой статьи в Biology Letters
Математические модели предсказывают, что в популяциях подверженных заражению паразитами организмов преимущество должны получить более редкие генотипы. В ходе экспериментов и наблюдений в природе уже были подтверждены многие следствия этого предсказания. Но только теперь в эксперименте на нематодах Caenorhabditis elegans и бактериях Serratia marcescens непосредственно продемонстрировано, что отбор на повышение вирулентности (смертности хозяев) приводит к тому, что паразиты становятся менее опасными для более редкого генотипа хозяина, тем самым давая ему селективное преимущество.
Изучение коэволюции паразитов и хозяев — хороший пример того, как в биологии переплетаются три метода исследований: математические модели, эксперименты и полевые наблюдения. Ни один из них не может дать окончательной картины без остальных, и у каждого есть свои преимущества и недостатки.
Именно из математических моделей в коэволюционную биологию пришло представление об отрицательном частотно-зависимом отборе как основной движущей силе коэволюции паразитов и хозяев. В этом варианте естественного отбора приспособленность того или иного генотипа (или фенотипа) отрицательно связана с частотой, то есть это «отбор в пользу редких вариантов». На качественном уровне эту идею изложил выдающийся британский биолог Джон Холдейн в своей статье-эссе «Болезни и эволюция» (J. B. S. Haldane, 1949. Disease and Evolution). Среди следствий этой идеи — и польза полового размножения в борьбе с паразитами (W. Hamilton, 1980. Sex versus non-sex versus parasite), и высокое и долго поддерживающееся разнообразие генов, задействованных в иммунитете (E. M. Leffler et al., 2013. Multiple Instances of Ancient Balancing Selection Shared Between Humans and Chimpanzees, см. также популярный рассказ об этой работе), и так называемый «осциллирующий полиморфизм» как один из основных вариантов антагонистической коэволюции. В этом случае генетический вариант (аллель или генотип) хозяина, ставший частым, вызывает адаптацию популяции паразитов (более успешными среди них окажутся те, кто способен эффективнее заражать частый генотип хозяина), что приведет к усилению паразитарной нагрузки, и, как следствие, к снижению приспособленности и уменьшению относительной частоты этого варианта в популяции. На смену прежде частому варианту придет другой, прежде редкий, который оказался менее подвержен заражению. А дальше история повторяется.
Случаи осциллирующего полиморфизма подтверждены в природе. Самый известный из них — это новозеландские улитки Potamopyrgos antipodarum и их паразиты-трематоды (группа плоских червей, также известная как сосальщики) Atriophallophorus winterbourni (M. F. Dybdahl, C. M. Lively, 1998. Host‐parasite coevolution: evidence for rare advantage and time‐lagged selection in a natural population). Интересно, что, хотя эти организмы уже более тридцати лет являются модельной системой для изучения коэволюции, вид сосальщиков был описан только в ноябре 2019 года (I. Blasco-Costa et al., 2019. A new species of Atriophallophorus Deblock & Rosé, 1964 (Trematoda: Microphallidae) described from in vitro-grown adults and metacercariae from Potamopyrgus antipodarum (Gray, 1843) (Mollusca: Tateidae)), а до этого он упоминался как Microphallus sp. Позднее в эксперименте на этой же модельной системе было показано, что за пять поколений популяция трематод эволюционирует в сторону лучшей способности заражать наиболее распространенный клон новозеландских улиток (B. Koskella, C. M. Lively, 2009. Evidence for negative frequency-dependent selection during experimental coevolution of a freshwater snail and a sterilizing trematode).
Еще один пример — это ржавчинный гриб Puccinia chondrillina и растение хондрилла ситниковая (Chondrilla juncea). В 13 из 16 популяций хондриллы в восточной Турции растения с наиболее частым генотипом были поражены паразитическим грибом, причем в 10 из них растения с другими генотипами от гриба вообще не страдали (P. Chaboudez, J. J. Burdon, 1995. Frequency-dependent selection in a wild plant-pathogen system).
Однако во всех этих случаях остается вопрос — дело именно в частоте данного генотипа, или в каких-то иных его свойствах? Чтобы ответить на этот вопрос, надо проверить, как поведут себя паразиты в популяциях хозяина, различающихся только частотой, но не набором генотипов. Именно такой эксперимент и был проведен Амандой Гибсон (Amanda Gibson) и командой исследователей из лаборатории Леви Морана (Levi Morran Lab) Департамента биологии Университета Эмори на модельной системе, состоящей из нематоды Caenorhabditis elegans и бактерии серратии Serratia marcescens.
Для опыта четыре изогенные линии C. elegans (обозначаемые N2, CB4856, LTM1 и ewIR68) смешивались попарно в соотношениях 9 к 1 и 3 к 4 (в дальнейшем анализе не учитывались различия в частоте, а только то, редким или частым был генотип). В ходе эксперимента паразиты в течение сотен поколений отбирались на большую вирулентность (смертоносность) для хозяина: к дальнейшему размножению допускались только бактерии из нематод, умерших в течение 24 часов после инфицирования. Таких раундов отбора в разных вариантах опыта было 10 или 20. Важно отметить, что каждый раз к бактериям, убивших нематод, добавлялись «наивные» черви из начальной популяции, то есть в данном случае возможность эволюционировать была дана только паразитам. Схема эксперимента представлена на рис. 1.
Сразу бросается в глаза невыровненность дизайна: разная длительность отбора, разная частота редких и частых генотипов. Дело в том, что представленные в обсуждаемой статье данные — это «побочный» продукт двух других экспериментов по изучению эволюции паразитов в гетерогенной популяции хозяев. Ранее в этих опытах было установлено, что используемые генотипы нематод различались по устойчивости к серратиям (рис. 2, слева), а используемый штамм серратии способен адаптироваться к каждому из изучаемых генотипов нематод в гомогенной популяции (рис. 2, справа), то есть среди хозяев нет сверхустойчивого клона.

Рис. 2. Слева — вирулентность исходного штамма Serratia marcescens по отношению к использованным в опыте генотипам нематод C. elegans: видно, что генотипы LTM1 и N2 менее устойчивы к этой бактерии. Справа — адаптация используемого штамма Serratia marcescens к генотипам нематод C. elegans в гомогенных популяциях. По вертикальной оси отложена разница в смертности хозяев после заражения бактериями, выращиваемыми с нематодами, и контролем, деленная на смертности бактерий из контроля. Контролем служили популяции бактерий, выращиваемых в отсутствии нематод и демонстрирующие изменение вирулентности за счет адаптации к иным, не связанным с наличием хозяев, условиям эксперимента. Эти результаты показывают, что используемый штамм серратий способен адаптироваться к используемым генотипам нематод, но в разной степени. Рисунки из дополнительных материалов к обсуждаемой статье в Biology Letters
В конце эксперимента смертность, вызываемую паразитами, оценивали на наивных нематодах тех же штаммов.
В двух из четырех пар генотипов хозяев серратии потеряли вирулентность по отношению к редкому генотипу, в одной — повысили вирулентность в отношении частого генотипа, и еще в одной — потеряли вирулентность по отношению и к редкому, и к частому генотипу (рис. 3). Последнее произошло с теми червями, которые исходно были менее чувствительны к бактериям.

Рис. 3. Изменение вирулентности (смертности хозяев) паразитов против редкого (rare) и частого (common) генотипа хозяина. По вертикальной оси отложена разность вирулентности после отбора и вирулентности предкового штамма паразита против данного генотипа хозяина, деленная на исходную вирулентность: положительные значения говорят об увеличении смертности, отрицательные — об уменьшении. Серые кружочки и линии отражают результат отдельных повторностей, цветные кружочки и черные линии — усредненные значения. Звездочками отмечены значимые различия. Рисунок из обсуждаемой статьи в Biology Letters
Статистический анализ показал, что совокупный результат эксперимента лучше всего описывается моделью, в которой вирулентность серратий понижается по отношению к редкому генотипу нематод на 3,73±1,74% и остается неизменной по отношению к частому генотипу (изменение 0,28±1,55%). При этом отсутствовало взаимодействие факторов «генотип» и «частота», то есть разные генотипы не различались по характеру изменения вирулентности в зависимости от частоты — не было такого, что, например, у одного генотипа падение вирулентности при редкой частоте оказывалось бы значимо большим, чем у другого. Этот результат был поддержан также двумя другими, независимыми статистическими проверками. Таким образом, можно утверждать, что в ходе этого эксперимента серратии теряли вирулентность по отношению к редкому генотипу, какой бы он ни был, и не меняли ее по отношению к частому генотипу.
Хотя полученный результат согласуется с тем, что наличие паразитов дает преимущество редким генотипам, размер эффекта довольно незначителен. В чем причины этого? Авторы отвергают два тривиальных объяснения малой эффективности отбора: недостаточная наследственная изменчивость у серратий (используемый штамм легко адаптируется к гомогенным популяциям нематод) и недостаточное время (наиболее сильные изменения произошли в вариантах опыта с 10, а не 20 раундами отбора). Два других предположения представляются авторам более правдоподобными. Во-первых, изучение средней вирулентности может скрывать внутрипопуляционные различия серратий, то есть отдельные линии последних могут проявлять особенно высокую вирулентность против частого или особенно малую вирулентность против редкого генотипа (замечу, впрочем, что исходная модель отрицательного частотно-зависимого отбора предполагает как раз изменение средней вирулентности по отношению к редкому и частому генотипу хозяина), — авторы вскользь упоминают, что у них есть подтверждающие такую изменчивость по вирулентности данные. Во-вторых, в этом эксперименте не было настоящей коэволюции, так как на каждом новом раунде отбора серратии сталкивались с наивными нематодами. А коэволюция, как неоднократно было показано, способствует генетической изменчивости у паразитов (см., например, R. D. Schulte et al., 2013. Host–parasite coevolution favours parasite genetic diversity and horizontal gene transfer), — авторы предполагают, что это может приводить к более сильным различиям между вирулентностью по отношению к редким и частым генотипам хозяев.
Несмотря на несовершенство дизайна, данный эксперимент наглядно продемонстрировал одно из основных положений коэволюционной биологии — наличие паразитов приводит к селективному преимуществу более редких генотипов (на всякий случай напомню, что пониженная смертность, вызываемая инфекцией, как раз и означает, что в данных условиях этот генотип оказывается более успешным).
Источник: elementy.ru





