













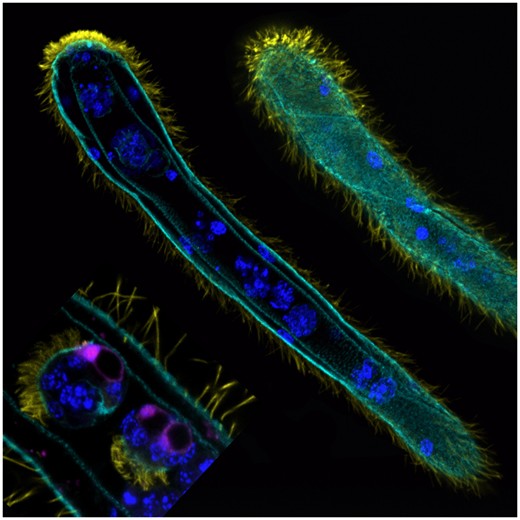

Рис. 1. Дициема на стадии ромбогена. Справа вверху — вид снаружи. В центре — вид в продольном разрезе. Слева внизу показана крупным планом часть ромбогена, в которой находятся маленькие многоклеточные инфузоригены — особи следующего поколения, которые проводят внутри осевой клетки ромбогена всю жизнь (на центральном изображении их тоже видно). Фото Tsai-Ming Lu из статьи C. McGrath, 2019. Highlight-Lessons on Parasitism from the Curious Dicyemida
Ортонектиды и дициемиды — две группы чрезвычайно просто устроенных паразитических животных, которых раньше считали переходными формами между одноклеточными и многоклеточными и объединяли в тип Mesozoa. Сейчас доказано, что простота ортонектид и дициемид — вторичная. Это сильно упростившиеся потомки каких-то двусторонне-симметричных животных из группы Spiralia, к которой относятся плоские и кольчатые черви. Насколько ортонектиды и дициемиды близки друг к другу — вопрос до сих пор дискуссионный. Но изучение их геномов, и в частности — серия недавно проведенных исследований геномов дициемид, в любом случае показывает, что их адаптивные стратегии были достаточно разными. Причем стратегия дициемид гораздо более необычна.
Слон и бактерия
Идет ли эволюция от простого к сложному?
Основатель эволюционной биологии Жан-Батист Ламарк был уверен, что да. Он считал, что по-другому и быть не может. Согласно ламарковскому принципу градации, тенденция к постоянному и неуклонному усложнению свойственна всему живому. На естественный вопрос «а почему же тогда до сих пор существуют очень простые организмы?» Ламарк отвечал: они позже возникли и просто еще не успели усложниться в такой мере, как, например, высшие животные. Принцип градации, таким образом, был тесно связан с гипотезой о многократном зарождении жизни.
Биология XX века убедительно показала, что эта гипотеза неверна. Вся жизнь на Земле имеет единое происхождение. Первым прямым свидетельством этого стало открытие в 1920-х годах биохимического механизма молочнокислого брожения: он оказался одним и тем же у бактерий и у млекопитающих. Серия подобных открытий позволила микробиологам сформулировать так называемую теорию биохимического единства жизни, кратко выраженную словами дельфтского профессора Альберта Яна Клюйвера (Albert Jan Kluyver): «От слона до маслянокислой бактерии — все одно и то же» (см. D. Singleton, R. Singleton, 2017. Remembering our forebears: Albert Jan Kluyver and the unity of life). Ну а последовавшая в 1960-х годах расшифровка генетического кода вообще сдала в архив все сомнения в том, что земная жизнь — это древо с единственным корнем. Все формы жизни, сколь угодно простые или сложные, должны найти на этом древе свое место (вернее, исследователи должны найти места, которые они там занимают: ведь эволюционное древо — это реальная структура, а не умозрительная модель). Что же касается сложности, то она по ходу ветвей древа в принципе может повышаться или понижаться — ни то, ни другое логически не запрещено.
Вверх и вниз по лестнице природы
В общем случае под сложностью системы можно понимать мощность набора параметров, которыми она определяется (Б. А. Старостин, 1974. Системный подход, параметры и сложность биологических объектов). Любая природная система в принципе может быть определена через сумму, во-первых, ее элементов и, во-вторых, связей между ними. Сложность любой данной системы тем выше, чем больше объем описания её элементов и их взаимодействий, минимально необходимого для понимания того, как она устроена (R. Hinegardner, J. Engelberg, 1983. Biological complexity).
Например, очевидно, что любая клетка сложнее единичной рибосомы. Это ясно хотя бы уже потому, что клетка содержит много рибосом — и, конечно, много других молекулярных структур, которые должны быть внутри нее более или менее закономерно расположены. Эукариотная клетка сложнее прокариотной, потому что она разделена мембранами на множество отсеков (компартментов), опять же расположенных относительно друг друга далеко не случайным образом. Атом железа сложнее атома водорода, потому что атом основного изотопа водорода (протия) состоит всего из двух элементарных частиц, а атом наиболее стабильного изотопа железа — из 82 частиц, если считать протоны, нейтроны и электроны. Ну и так далее.
В качестве грубого, но удобного критерия сложности многоклеточного организма американский биолог Джеймс Валентайн (James Valentine) предложил использовать количество морфологически различимых типов клеток (морфологически различимых — значит, попросту говоря, таких, которые можно отличить друг от друга под обычным световым микроскопом). Действительно, все типы клеток — покровные, железистые, нервные, мышечные и какие угодно еще — существуют не просто так, а потому, что у каждого из них есть какая-то физиологическая функция. Поэтому полный список типов клеток емко и в то же время достаточно содержательно характеризует устройство организма как целого. Валентайн дает единообразную для всех животных нормированную оценку: например, у стрекающих число типов клеток равно примерно 10, у нематод их 14, у коловраток — 15, у гребневиков — 17, у приапулид — 20, у полухордовых — 25, у кольчатых червей и моллюсков 37–38, у иглокожих — 41, у всевозможных членистоногих — от 42 до 90, а у человека — 210 (J. W. Valentine, 2003. Cell types, numbers, and body plan complexity). У трихоплакса, одного из самых простых многоклеточных животных, типов клеток по тому же подсчету всего 4.
На протяжении истории Земли сложность животных организмов в целом возрастала (Валентайн соорудил серию графиков, подтверждающих это интуитивное впечатление; см. J. W. Valentine, 1996. The evolution of complexity in metazoans). Но ясно, что этот процесс, во-первых, неравномерен, и во-вторых — не однонаправлен. В некоторых эволюционных ветвях сложность уменьшается. Ничего удивительного в этом нет. Сложность — такой же признак, как и любое другое свойство организма, так же подверженный действию естественного отбора и других факторов эволюции. Сам Валентайн упоминает в своих сводках как минимум три группы животных, у которых совершенно точно произошло сильное вторичное упрощение, то есть уменьшение сложности. Это миксозои (3 типа клеток), ортонектиды (3 типа клеток) и дициемиды (4 типа клеток). Однако сейчас мы понимаем, что эволюционные судьбы этих трех групп во многом различны.
Примечательнее всего обстоит дело с миксозоями. Это микроскопические паразиты, у которых питающаяся стадия представляет собой, по сути, просто многоядерную амебу, а расселительная стадия — многоклеточную спору (чаще всего она состоит из 6 клеток: две клетки-зародыша, две клетки-створки и две стрекательные клетки). На многоклеточных животных миксозои совершенно не похожи. В старых учебниках их вообще относили к простейшим. Однако в 1990-х годах молекулярные и электронно-микроскопические исследования показали, что миксозои — это на самом деле сильно упрощенные потомки стрекающих (группа, в которую входят кораллы, гидры, медузы и тому подобные существа). У стрекающих есть эпителии, нервная система, мускулатура — в общем, продвинутая тканевая организация, которую миксозои совершенно утратили.
Около года назад российские биоинформатики опубликовали удивительную гипотезу: предполагается, что миксозои произошли не от «нормального» упрощенного паразитического животного, а от клеток трансмиссивной раковой опухоли (A. Y. Panchin et al., 2019. From tumors to species: a SCANDAL hypothesis; см. также научно-популярный обзор в блоге одного из авторов работы — Александра Панчина). Инфекционный рак — вообще любопытное явление, и неудивительно, что в последнее время он стал популярной темой исследований (см. М. С. Гельфанд, 2016. Инфекционный рак). В данном случае под инфекционным раком подразумеваются не опухоли, вызываемые вирусами (хотя в принципе возможны и такие), а опухоли, распространяемые самими раковыми клетками, которые переходят с одной особи животного на другую, как обычные микроорганизмы. Широко известные примеры таких опухолей — трансмиссивная венерическая саркома собак и лицевая опухоль сумчатого дьявола. Первая не слишком опасна (она легко излечивается), а вот вторая имеет злокачественный характер, и ее эпидемия всерьез угрожает существованию сумчатого дьявола как вида. Кроме того, несколько примеров трансмиссивных раковых опухолей обнаружено почему-то у двустворчатых моллюсков (см. Моллюски разных видов могут заражать друг друга раком, «Элементы», 27.10.2016). Такая опухоль представляет собой самостоятельный колониальный организм — не хуже какой-нибудь колонии водорослей. Спрашивается, почему бы на ее основе не развиться новым эволюционным ветвям?
В пользу этой гипотезы приведено несколько серьезных аргументов. Во-первых, у большинства миксозоев никакие стадии жизненных циклов не имеют ничего общего с нормальным эмбриональным развитием многоклеточных животных. У них нет ни бластулы, ни гаструлы, ни зачатков каких-либо органов. Более того, самая сложная стадия развития типичных миксозоев — многоклеточная спора — состоит из гаплоидных клеток (с одним набором хромосом), что для многоклеточных животных в высшей степени необычно: как правило, у них гаплоидны только яйцеклетки и сперматозоиды (см., например: A. V. Uspenskaya, 1982. New data on the life cycle and biology of Myxosporidia). Во-вторых, внимательный анализ генома показал, что у миксозоев отсутствуют некоторые белки (например, каспазы), обычно участвующие в процессе апоптоза, то есть программируемой гибели ненужных клеток. Эти белки есть у всех других животных, включая губок, трихоплакса и любых паразитов. А вот в раковых клетках они исчезают (вместе с соответствующими генами). Все это позволяет допустить, что миксозои в самом буквальном смысле произошли от раковых клеток, которые научились жить самостоятельно и вторично выстроили некое подобие многоклеточного организма. Такой способ видообразования, конечно, необычен, но ничего невозможного в нем нет.
Проблема в том, что класс миксозоев состоит из двух подклассов: миксоспоридии, к которым в полной мере относятся все перечисленные аргументы, и малакоспоридии, которые устроены гораздо сложнее (см. E. Jimenez-Guri et al., 2007. Buddenbrockia is a cnidarian worm). У малакоспоридий есть нормальное (хоть и упрощенное) эмбриональное развитие, вполне себе двусторонне-симметричное червеобразное тело и даже мышечные клетки (A. Gruhl, B. Okamura, 2012. Development and myogenesis of the vermiform Buddenbrockia (Myxozoa) and implications for cnidarian body plan evolution). О том, как выглядел общий предок всех миксозоев и каковы были начальные этапы их эволюции, наверняка смогут поведать геномы малакоспоридий, но пока они еще не секвенированы. Поэтому гипотеза о происхождении миксозоев от раковой опухоли остается пока не более чем гипотезой. Но в любом случае принципиальную возможность такого способа эволюции стоит иметь в виду.
Что же касается непосредственно миксоспоридий, то они, несомненно, достигли самого крайнего упрощения, какое возможно для животных. Следующим шагом в эту сторону могла бы быть только полная потеря многоклеточности. Авторы упомянутой статьи обоснованно называют такое упрощение катастрофическим.
Со второй группой сильно упрощенных паразитов — ортонектидами — дело обстоит совсем иначе (см. Одни очень простые животные паразитируют внутри других: ортонектида в ксенотурбелле, «Элементы», 13.02.2019). Раньше многие зоологи считали ортонектид переходными формами между одноклеточными и многоклеточными, и только электронно-микроскопические исследования показали, что их простота была преувеличена: на самом деле у ортонектид есть нервные и мышечные клетки, и в целом они сохраняют план строения своих предков — кольчатых червей (а то, что ортонектиды произошли от кольчатых, более или менее надежно установлено молекулярной филогенетикой). Правда, такие системы, как пищеварительная и выделительная, у них исчезли бесследно, да и геном сильно уменьшен по сравнению с «нормальными» червями.
Загадочные дициемиды
Дициемид открыл еще в XVIII веке профессор зоологии Неаполитанского университета Филиппо Каволини (Filippo Cavolini). Это микроскопические червеобразные существа, живущие в телах головоногих моллюсков и обладающие поразительно сложными жизненными циклами. По словам Джеральда Даррелла, если Фрейд полагал, что половая жизнь средней человеческой особи очень сложна, то он получил бы нервное расстройство, доведись ему подвергнуть психоанализу синеголового губана (Thalassoma bifasciatum); уже то, что один и тот же экземпляр этой рыбы может в разные периоды жизни быть самцом и самкой, заставило бы великого психоаналитика призадуматься. Но у дициемид все еще более необычно. Во-первых, у них (как у многих паразитов) множество бесполых поколений чередуется с регулярно возникающими половыми. Во-вторых, их жизненный цикл включает не менее пяти самостоятельных многоклеточных стадий, по-разному устроенных и имеющих собственные названия, причем — в отличие от нормальных червей — тут не так-то просто решить, какую из этих стадий следует считать главной («взрослой»). В-третьих, очень характерная особенность дициемид состоит в том, что при смене поколений у них одни клетки образуются внутри других. И не только клетки, но и зародыши, и целые организмы. В жизненном цикле дициемид есть многоклеточная форма, которая в норме проводит всю свою жизнь не просто внутри другой многоклеточной формы (предыдущего поколения), а внутри одной из ее клеток (рис. 1). Такой ход развития крайне необычен для кого угодно, и для животных — уж точно.
В отличие от ортонектид, которые паразитируют на самых разных морских беспозвоночных, дициемиды живут в почках головоногих моллюсков и только там. Никаких промежуточных хозяев у них нет. Зато многие виды осьминогов и каракатиц заражены в природе дициемидами на 100%. Особого вреда своим хозяевам дициемиды, судя по всему, не наносят — они поглощают те питательные вещества, которые удается найти в моче. Причудливый жизненный цикл дициемид подробно описан в книге Владимира Васильевича Малахова «Загадочные группы морских беспозвоночных», куда и можно обратиться за деталями (рис. 2).

Рис. 2. Жизненный цикл дициемид. Все многоклеточные стадии, кроме инфузориформа, проводят всю жизнь внутри тела головоногого моллюска. Одна из стадий — инфузориген — живет в цитоплазме осевой клетки ромбогена; это показано также на рис. 1. Иллюстрация из книги В. В. Малахова «Загадочные группы морских беспозвоночных»
Главная питающаяся стадия дициемид является бесполой и называется нематогеном (буквально это значит что-то вроде «рождающий червей»). Нематоген состоит из единственной очень длинной осевой клетки (именно внутри нее развиваются зародыши следующего поколения), со всех сторон окруженной покровными клетками, которые несут реснички. Этих покровных клеток бывает 20–30, причем их точное число и расположение строго постоянно для каждого вида дициемид. Никакой полости тела у дициемид нет, и никаких остатков нервной и мышечной систем у них (в отличие от ортонектид) не нашли. Скорее всего, и не найдут. Дело в том, что дициемидам — как и ряду других мелких многоклеточных животных — свойственна эвтелия, то есть постоянство клеточного состава. Это означает, что окончательное число клеток, одинаковое для всех особей данного вида, достигается в конце эмбрионального развития и больше не меняется. Нематоген дициемид может расти, но никаких клеточных делений в нем при этом не происходит (если не считать размножения). В эвтелическом организме каждая клетка на счету, и каждая занимает свое уникальное, раз и навсегда определенное место. Да и клеток в теле дициемид считанные десятки, а не сотни, как у ортонектид. Это понятно: малое число клеток — обычный спутник эвтелии. Вряд ли при таких условиях исследователи могли бы не заметить у дициемид нервные или мышечные клетки. Вероятно, их просто нет. Значит, дициемиды устроены совсем иначе, чем ортонектиды, с которыми их традиционно относили к одному типу.
Откуда же эти удивительные существа взялись?
Палеонтологическая летопись ответить на этот вопрос не поможет: дициемиды — не те организмы, которым легко в ней сохраниться. Но остается возможность прочитать геномы современных видов дициемид, чтобы извлечь заключенные там сведения об эволюционном пути этих животных, а заодно прояснить и их нынешнее устройство.
В 2019 году почти одновременно вышли две статьи с анализом геномов дициемид. Одну опубликовала группа китайских и японских авторов, среди которых значится известный биолог Нориюки Сато (Noriyuki Satoh). Другую — международная группа исследователей, один из которых (Леонид Леонидович Мороз, известный своими исследованиями гребневиков; см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015) работает преимущественно в США, а остальные — в Москве. Приятно отметить, что восемь из одиннадцати авторов этой второй статьи — сотрудники МГУ. Двое из них, доктора Владимир Вениаминович Алешин и Юрий Валентинович Панчин, участвовали и в обсуждаемой выше работе, посвященной миксозоям. Выводы, сделанные в двух статьях про дициемид, отлично дополняют друг друга, хотя кое-где между ними есть и разногласия. Тем интереснее.
Сумрачный лес деревьев спиралий
Занимаясь любой группой животных, биологи всегда стремятся для начала выяснить, каково ее место на филогенетическом древе. Иными словами, от кого она произошла (ну хотя бы предположительно), кто ее близкие родственники, а кто дальние. Для современных организмов, у которых доступен геном, молекулярные методы, как правило, позволяют определять родство довольно точно, просто за счет того, что они оперируют огромными объемами информации.
Но и молекулярная филогенетика иногда буксует — например, если эволюция генома в интересующей нас группе шла слишком быстро. В этом случае множественные и повторные мутации могут полностью стирать информацию о родстве, давая взамен ложный сигнал (см. В. В. Алешин и др., 2010. Эволюция предельно упрощенных животных — ортонектид). Для ортонектид и дициемид эта проблема как раз актуальна: и у тех, и у других эволюция, видимо, шла в какие-то отрезки времени чрезвычайно быстро, и поэтому вычленить из их геномов филогенетически значимую информацию оказалось очень трудно. В отношении ортонектид молекулярным филогенетикам в конце концов удалось в первом приближении прийти к согласию: похоже, что эта ветвь находится внутри группы кольчатых червей. Другими словами, ортонектиды — прямые потомки кольчатых.
С дициемидами все гораздо сложнее. Сейчас можно уверенно утверждать, что они, как и ортонектиды, относятся к эволюционной ветви под названием Spiralia. Проблема в том, что эта ветвь поистине огромна, и родственные отношения внутри нее запутаны, пожалуй, сильнее, чем в любой другой крупной группе животных (см. Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016). Кроме ортонектид и дициемид, в группу спиралий входят кольчатые черви, моллюски, брахиоподы, плоские черви, коловратки, мшанки, немертины и еще несколько типов. И вот положение дициемид внутри спиралий — что неудивительно — до сих пор остается темой споров. Именно здесь выводы группы Сато расходятся с выводами группы Алешина. Пару лет назад группа Сато считала, что дициемиды вместе с ортонектидами образуют ветвь, родственную плоским и брюхоресничным червям (T. M. Lu et al., 2017. The phylogenetic position of dicyemid mesozoans offers insights into spiralian evolution). В новой работе они, хотя и не обсуждают филогенетическое положение дициемид подробно, держатся того же мнения и особенно подчеркивают, что ортонектиды с дициемидами — ближайшие родственники. А это, в свою очередь, позволяет сохранить старинное объединение этих двух групп под названием Mesozoa (букв. «среднеживущие»; это название появилось, когда ортонектид и дициемид считали переходными формами между одноклеточными и многоклеточными животными).


Рис. 3. Эволюционное древо Spiralia, построенное группой Алешина. Здесь ортонектиды и дициемиды находятся далеко друг от друга, никаких оснований объединять их в одну группу нет. Иллюстрация из обсуждаемой статьи в Frontiers in Genetics
Секвенировав геном и преодолев множество вычислительных трудностей, группа Алешина пришла к выводу, что ортонектиды на эволюционном древе спиралий уверенно группируются с кольчатым червями, а вот дициемиды занимают там совсем другое положение — очень изолированное и неустойчивое. На разных вариантах древес ветвь дициемид оказывается на разных местах, но неизменно далеко от кольчатых (ближе к плоским и брюхоресничным или даже к таким малоизвестным группам, как внутрипорошицевые и циклиофоры). Есть и версии деревьев, где дициемиды оказываются сестринской группой ко всей огромной ветви Lophotrochozoa, в которую входят кольчатые черви, моллюски, немертины, брахиоподы, мшанки и форониды. Но ни в одном из этих вариантов положение дициемид не имеет ничего общего с положением ортонектид. Алешин и его коллеги особенно подчеркивают этот последний момент. Если они правы, то никакого типа мезозоев не существует. Ортонектиды и дициемиды не связаны ничем, кроме того, что и те и другие — спиралии. Это разные эволюционные ветви, лишь отдаленно родственные друг другу и совершенно независимо пошедшие по пути специализации к паразитизму.
Что же с ними на этом пути случилось?
Тайны генома дициемид
Исследования прочитанных геномов дициемид выявили там довольно много признаков, свидетельствующих о бурных эволюционных событиях. Перечислим некоторые из них.
1. Маленький размер генома. Ясно, что размер генома, то есть количество находящейся в клетке ДНК — показатель, важный для любого организма (хотя интерпретировать его бывает непросто; см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Размер генома изученного представителя дициемид — он называется дициемой — равен 67,5 мегабаз. Что это значит? Мегабаза — это общепринятая единица измерения количества ДНК: миллион пар азотистых оснований или миллион пар нуклеотидов (в данном случае это одно и то же). Шестьдесят семь мегабаз — много это или мало? Это мало. Например, у мухи дрозофилы геном имеет размер около 180 мегабаз, у ланцетника — примерно 520 мегабаз, а у человека — больше 3 тысяч мегабаз. Для многоклеточного животного геном дициемы крайне невелик.
В результате чего он стал таким маленьким? Начнем с того, что на этот вопрос в принципе возможны разные ответы. Причиной уменьшения генома может быть уменьшение числа рабочих генов, а может быть и уменьшение суммарного количества ДНК за счет некодирующих — то есть, грубо говоря, бессмысленных — нуклеотидных последовательностей. Судя по всему, в эволюции дициемид происходило и то, и другое. У дициемы обнаружено 5012 белок-кодирующих генов — очень мало по животным меркам (для сравнения: у дрозофилы насчитывается 14 тысяч белок-кодирующих генов, а у человека — примерно 20 тысяч). Уменьшение числа белок-кодирующих генов, конечно же, отражает упрощение функций организма. Но и средний размер интронов, то есть некодирующих внутригенных последовательностей, у дициемы тоже оказался сильно уменьшен по сравнению с большинством животных — а это верный знак, что одной из движущих сил редукции генома тут было именно уменьшение общего количества ДНК. А зачем может быть нужно уменьшать количество ДНК? Да, например, затем, чтобы клетки быстрее делились. Хотя и не факт, что эта причина единственная.
Связано ли уменьшение генома дициемид с паразитизмом? Вероятнее всего, да. Однако связь тут непростая и нелинейная. В пользу объяснения через паразитизм говорит тот факт, что подобное уменьшение генома отмечено у ленточных червей и у обсуждавшихся выше ортонектид, причем, как и у дициемид, в этих группах одновременно наблюдается падение числа генов и уменьшение среднего размера интронов. Есть, правда, и контрпример — кровяная двуустка, очень специализированный паразит из группы сосальщиков, у которого геном тем не менее относительно большой (363 мегабазы; M. Berriman et al., 2009. The genome of the blood fluke Schistosoma mansoni). С другой стороны, один из самых маленьких геномов, какие вообще есть у животных — 70 мегабаз — обнаружен у аппендикулярий, плавающих в море низших хордовых, которые никогда паразитами не были. Так что в причинах уменьшения генома надо еще разбираться: этот параметр наверняка зависит от нескольких разных факторов.
2. Небольшое число Hox-генов. Как известно, гены семейства Hox играют важную роль в развитии двусторонне-симметричных животных, регулируя дифференцировку отделов тела и их отличия друг от друга (см., например: Программы работы Hox-генов у личинок и взрослых особей кольчатого червя принципиально отличаются, «Элементы», 27.05.2013). Это генное семейство довольно велико: например, у морского кольчатого червя нереиса 11 Hox-генов, у ланцетника — 15, у человека — 39, а у некоторых других позвоночных бывает и больше. А у дициемы всего три Hox-гена. Примечательно не только их малое число, но и то, что у дициемы не обнаружены так называемые «передние» Hox-гены, которые у более сложных животных экспрессируются в передней части тела и контролируют развитие головы. Это означает, что у дициемид обрушилась важнейшая для двусторонне-симметричных животных часть плана строения. Ну а действительно, зачем им голова?
У изученного по этому признаку представителя ортонектид Hox-генов тоже всего три. Но состав их другой: один относится к «передней» группе и два — к «центральной» (K. Mikhailov et al., 2016. The genome of Intoshia linei affirms orthonectids as highly simplified spiralians). Сохранение «переднего» гена объяснить легко: в отличие от дициемид, у ортонектид остается пусть и очень простая, но все-таки действующая нервная система, а для ее развития экспрессия Hox-генов обычно крайне важна, и особенно — в передней части тела. Кроме того, у ортонектид есть как минимум один ParaHox-ген. Генное семейство ParaHox невелико, родственно семейству Hox и присуще подавляющему большинству животных (см. Дупликация гомеобоксных генов могла быть одной из причин кембрийского взрыва, «Элементы», 13.02.2015). У дициемид ParaHox-гены вообще не найдены, и это должно означать, что они там вторично исчезли. Все эти факты подтверждают, что упрощение плана строения у ортонектид и дициемид шло по-разному, а значит — независимо.
С другой стороны, один (а может быть, и не один) из Hox-генов сохранившейся «центральной» группы у дициемид неожиданно дуплицировался, то есть породил собственную работоспособную копию. Зачем — пока неизвестно. Но это наглядно показывает, что к прямолинейному упрощению эволюция дициемид не сводится.
3. Отсутствие базальных мембран. Характерной чертой животных, отличающей их от других эукариот, можно назвать обилие эпителиев — сплошных слоев клеток, объединенных специальными контактами и как бы наклеенных на пленку, которая называется базальной мембраной. Эпителии подвижны, они могут выгибаться, впячиваться, образовывать полости и трубки, создавая в результате форму животного организма. Ни у кого, кроме животных, нет ничего подобного (правда, аналоги эпителиев бывают в плодовых телах слизевиков, но никаких формообразовательных движений они там не совершают). Базальная мембрана — важнейшая принадлежность эпителия, его механическая основа, — состоит из нескольких структурных белков, главные из которых — коллаген IV типа и ламинин. Выяснилось, что у дициемид просто нет генов, кодирующих эти белки. Таким образом, строить базальные мембраны им не из чего. Отказ от полноценных эпителиев — событие, редкое в животном мире, а для спиралий вообще уникальное. Это очень высокая степень упрощения организма.
4. Отсутствие потенциал-зависимых натриевых каналов. Любой ионный канал — это не что иное, как очень сложная белковая молекула, встроенная в клеточную мембрану и избирательно пропускающая сквозь нее какие-нибудь ионы (натрия, калия, хлора, кальция или водорода; см. Эволюция ионных каналов шла у животных параллельно, «Элементы», 05.03.2015). Особое значение для животных имеет разновидность ионных каналов, которая называется потенциал-зависимыми натриевыми каналами. Именно они запускают механизм возбуждения нервной клетки. Без потенциал-зависимых натриевых каналов проведение нервного импульса физически невозможно. И вот у дициемид генов, кодирующих эти каналы, не оказалось. От остатков нервной системы они отказались полностью.
Или все-таки не полностью? Алешин с коллегами обращают внимание на то, что у дициемид сохранились белковые рецепторы к глутамату, причем разных типов: как метаботропные, действующие через обмен веществ, так и ионотропные — связанные с ионными каналами. Глутамат — это очень важный для большинства животных нейромедиатор, то есть вещество, передающее сигналы от одной нервной клетки к другой. Более того, у дициемид, по-видимому, есть большая группа ионотропных рецепторов к еще более важному нейромедиатору — ацетилхолину. Что конкретно делают все эти рецепторы в организме, лишенном нервной системы, пока не выяснено. Но Алешин с коллегами высказали по этому поводу очень интересную гипотезу. У дициемид, конечно, нет натриевых ионных каналов, но зато совершенно точно есть кальциевые. Это неудивительно: кальциевая проводимость — гораздо более древняя и универсальная, хотя и медленная. На ее основе вполне можно создать систему межклеточной сигнализации, аналогичную нервной. Что-то в этом роде есть у губок (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Такая система должна быть гораздо медленнее и примитивнее настоящей нервной, но существам вроде дициемид ее вполне хватит.

Рис. 4. Гипотетическая модель межклеточной сигнализации у дициемид, построенная группой Алешина исключительно по генетическим данным. АЦХ — ацетилхолин. Любой читатель, знакомый с физиологией, сразу увидит, что эта конструкция во многом похожа на обыкновенный нервно-мышечный синапс… за единственным исключением: здесь ни в одном процессе не участвует натрий. Иллюстрация из обсуждаемой статьи в Frontiers in Genetics
5. Разрастание отдельных групп генов на фоне общей редукции. Безусловно, геном дициемид очень сильно упрощен. Но, как уже сказано, его эволюция к этому не сводится. Усложнение там тоже происходит — во всяком случае, в некоторых отдельных областях. Есть гены, которые у дициемид удвоились, а потом еще удвоились, породив в итоге целые родственные кластеры (до шести генов, когда в принципе могло бы хватить и одного). Очевидно, так ведут себя гены, продукты которых особенно важны для организма, в данном случае — для организма специализированного паразита. Например, это гены, кодирующие белки, которые обеспечивают эндоцитоз — захват отдельной клеткой внешнего материала посредством впячивания мембраны и ее замыкания в пузырьки, уходящие внутрь. Ясно, что для паразита, у которого нет никакой пищеварительной системы, это основной способ питания. В эндоцитозе всегда принимают участие микрофиламенты — внутриклеточные сократимые нити, состоящие из белка актина, поэтому число генов актина у дициемы тоже увеличено. То же самое относится к генам динеинов, моторных белков, которые обеспечивают перемещение по клетке мембранных пузырьков, возникших в результате эндоцитоза и содержащих питательные вещества.
6. Необычный митохондриальный геном. Широко известно, что митохондрии, органеллы эукариотных клеток, обеспечивающие кислородное дыхание, — это бывшие бактерии. Они до сих пор размножаются делением, и неудивительно, что в них есть собственная ДНК. Чаще всего это просто кольцевая ДНК бактериального типа, несущая некоторое небольшое число генов (например, у человека митохондриальных генов 13, если считать только белок-кодирующие). У дициемы ситуация другая. Ее митохондриальный геном состоит из множества маленьких кольцевых ДНК, которые, как правило, несут всего по одному гену, и только в нескольких случаях — по два. Такая фрагментация митохондриального генома — вещь редкая, хотя и не уникальная: она отмечена у картофельных нематод и у вшей (R. Shao et al., 2009. The single mitochondrial chromosome typical of animals has evolved into 18 minichromosomes in the human body louse, Pediculus humanus). Можно обратить внимание, что и картофельные нематоды, и вши, и дициемиды — быстро эволюционирующие паразиты. Но, какова бы ни была здесь причинная связь, сейчас важнее отметить, что по этому признаку дициемиды совсем не похожи на ортонектид. Митохондриальный геном ортонектид — традиционный, с одной большой кольцевой ДНК.
Мозаика эманации
Упрощение структуры организма — процесс, давно знакомый биологам-эволюционистам. Академик Алексей Николаевич Северцов называл его дегенерацией, а ученик Северцова академик Иван Иванович Шмальгаузен — катаморфозом. Современный палеонтолог Александр Сергеевич Раутиан, ученик учеников Шмальгаузена (эта цепочка продолжается), предложил более общую систему понятий (А. С. Раутиан, 1988. Палеонтология как источник сведений о закономерностях и факторах эволюции). Существует два режима развития: специализация, когда скорость запечатления в структуре новой информации (запоминания) превосходит скорость потери информации (забвения), и эманация, когда дело обстоит наоборот. Всевозможные дегенерации и катаморфозы — частные случаи эманации.
Как правило, в ходе эманации происходит не только уменьшение числа элементов системы, но и понижение мощности коррелятивных (то есть функциональных) связей между оставшимися элементами. Проще говоря, эманирующая система становится менее целостной. Очевидно, например, что колония клеток, ведущая образ жизни трансмиссивной раковой опухоли — менее целостная система, чем организм млекопитающего, от которого она произошла. Именно поэтому при эманации должен быть чрезвычайно силен эффект мозаичности (А. Л. Тахтаджян, 1946. Об эволюционной гетерохронии признаков), когда разные «функциональные блоки» организма эволюционируют с разными скоростями и в разных направлениях, и все это по-разному комбинируется в разных эволюционных ветвях. Если существует несколько параллельно эманирующих ветвей, пусть даже и происходящих от близких предков, следовало бы из этих соображений ожидать, что их адаптивные стратегии окажутся разными и порядок, в котором приобретаются и теряются признаки, будет сильно отличаться.
Именно это мы и видим, когда сравниваем ортонектид и дициемид. Примеры приведены выше, а сейчас можно добавить еще один. В мышечных клетках двусторонне-симметричных животных есть белок тропонин. Он характерен для поперечнополосатых мышц и для сердечной мышцы, а в гладких мышцах отсутствует. У ортонектид никаких тропонинов нет, гены соответствующих белков утрачены: это понятно, хотя мышечные клетки у них и имеются, но только гладкие. А вот у дициемид, которые вообще отказались от любых мышц, тропонин сохранился! Что он там делает, мы пока точно не знаем, но скорее всего — принимает участие в медленном сокращении покровных или осевых клеток. Это типичный «перекрест направлений специализации», если воспользоваться терминологией академика Армена Леоновича Тахтаджяна.
Кроме того, на примерах ортонектид и дициемид хорошо видно, что эманация никогда не бывает равномерной. Пусть даже большинство частей системы упрощается, но какие-то отдельные звенья обязательно будут совершенствоваться, а скорее всего и усложняться. Это — та необходимая специализация, без которой вообще нет ни эволюции, ни выживания. «Жить — значит специализироваться».
Ортонектиды и дицемиды входят в группу Spiralia, но если происхождение ортонектид по обсуждаемой версии связано с кольчатыми червями, то откуда взялись дициемиды — все еще неясно. Есть старая гипотеза, что происхождение дициемид связано с паразитическими плоскими червями из группы сосальщиков, которые похожи на них некоторыми особенностями жизненного цикла (Т. А. Гинецинская, 1968. Трематоды, их жизненные циклы, биология и эволюция). Не исключено, что это верно. Но установление точных родственных связей дициемид — пока что дело будущего.
Источник: elementy.ru





