













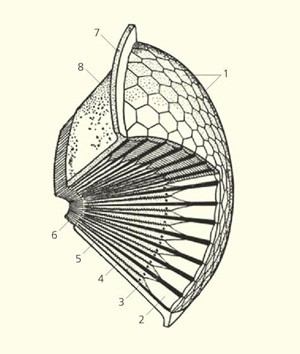


Рис. 1. Схема строения фасеточного глаза: 1 — роговичные линзы, 2 — кристаллический конус, 3 — пигментные клетки, 4 — ретинальные клетки, 5 — рабдом, 6 — аксоны ретинальных клеток, 7 — покровы головы, 8 — глазная капсула. Рисунок из статьи А. Сергеев, А. Благодатский, 2015. Насекомые и бионика: загадки зрительного аппарата
Немецкие палеонтологи тщательно исследовали строение фасеточного глаза трилобита Aulacopleura koninckii возрастом 429 млн лет. Уникальная сохранность остатков позволила детально рассмотреть и изучить строение отдельных омматидиев и глазных капсул. Оказалось, что по своему устройству глаза A. koninckii очень похожи на глаза современных насекомых. Древнейшими известными обладателями фасеточных глаз были кембрийские трилобиты Schmidtiellus reetae, жившие более 500 млн лет назад, причем их глаза были устроены крайне примитивно. Новая находка указывает на то, что эволюция зрительных органов трилобитов шла довольно быстро и в том же направлении, что и у других членистоногих.
Фасеточные глаза характерны для насекомых, ракообразных и некоторых других членистоногих. Почему эти животные «выбрали» такой принцип устройства зрительного аппарата, который не встречается больше ни у кого? Он имеет свои преимущества. Во-первых, хотя фасеточный глаз и занимает большую площадь, он относительно неглубоко уходит внутрь головы насекомого и тем самым экономит там место. Во-вторых, уступая в остроте зрения глазам позвоночных, он позволяет гораздо лучше видеть быстро движущиеся объекты. Наконец, у некоторых насекомых фасеточные глаза различают степень поляризации света — очень полезная способность.
Общий принцип устройства фасеточного глаза таков (рис. 1). Он сложен из маленьких конусовидных глазков, которые называют фасетками или омматидиями. Их число сильно варьирует: от нескольких штук у щетинохвосток до 30 тысяч у стрекоз. Наружная стенка омматидия — роговица, шестигранная двояковыпуклая линза из хитина. Под ней расположена вторая линза — прозрачный кристаллический конус. Преломленный ими свет попадает на цилиндрический пучок из (обычно восьми) длинных зрительных ретинальных клеток. Часть каждой из этих клеток, обращенная внутрь цилиндра, называется рабдомером, в ней содержится зрительный пигмент родопсин. Вместе рабдомеры в каждом цилиндре образуют зрительную палочку — рабдом. Она и воспринимает свет, аналогично нашей сетчатке.
По бокам омматидий окружают клетки, содержащие черный пигмент, который поглощает световые лучи. Благодаря им на рабдом каждого омматидия попадает только свет, преломленный его линзами, но не боковой свет из других омматидиев. Впрочем, так только в аппозиционных глазах насекомых, приспособленных к яркому дневному свету. У ночных же и сумеречных насекомых изображения, поступающие от соседних омматидиев, могут суммироваться, что позволяет им лучше видеть в темноте. Это достигается либо за счет перераспределения черного пигмента в светоизолирующих клетках, либо уже с помощью объединения сигналов нервной системой. Такой тип фасеточного глаза называется суперпозиционным.
Как давно появились фасеточные глаза? Первыми известными их обладателями были трилобиты, примитивные морские членистоногие, которые появились на заре палеозоя, в кембрийском периоде. Самым древним из пока что известных является фасеточный глаз трилобита вида Schmidtiellus reetae, жившего свыше 500 млн лет назад — его описали по окаменелости в 2017 году профессор Бригитте Шонеманн (Brigitte Schoenemann) из Кельнского университета и ее коллеги (B. Schoenemann et al., 2017. Structure and function of a compound eye, more than half a billion years old).
Омматидиев в фасеточном глазе S. reetae было всего около 100. Их роговица еще не превратилась в линзу, а была просто плоской полупрозрачной кутикулой, неспособной фокусировать свет. Каждый омматидий были «упакован» в отдельную «корзинку» из клеток кутикулы. Однако пигментных клеток вокруг них еще не было, так что свет мог попадать из одного соседнего омматидия в другой. В отличие от современных ночных форм с суперпозиционными глазами, это было не приспособлением, а примитивной чертой — фасеточный глаз S. reetae оставался аппозиционным и обеспечивал своему обладателю, по совокупности черт, довольно плохое зрение.
Теперь та же команда ученых опубликовала в журнале Scientific Reports новую статью, с описанием фасеточного глаза более «молодого» трилобита Aulacopleura koninckii. Окаменелые остатки A. koninckii возрастом 429 млн лет, найденные в Чехии, содержат, в том числе, один хорошо сохранившийся фасеточный глаз — и его изучение показало, что он уже мало чем отличается от глаз современных насекомых и ракообразных.
В глазе A. koninckii было уже около 200 фасеток, некоторые из которых авторы статьи, благодаря хорошей сохранности экземпляра, смогли разглядеть в цифровой микроскоп достаточно хорошо. Оказалось, что, как у большинства современных форм, в фасетках этого трилобита были пучки из восьми ретинальных клеток. На рис. 2 они показаны их поперечные срезы, и можно разглядеть характерные розетки с рабдомом в центре. На некоторых срезах видно, что рабдом окружен серой каймой. По-видимому, это остатки так называемого «палисада» — внеклеточной области с низким показателем преломления, которая призвана улучшить восприятие света. Для этой же цели у самого рабдома был сравнительно большой диаметр (около 8 мкм).

Рис. 2. Окаменелые остатки Aulacopleura koninckii, ставшие материалом обсуждаемого исследования. a — общий вид; b — рисунок-реконструкция внешнего вида; c — срез всей окаменелости; d — срез головной части (цефалона); e — сохранившийся левый фасеточный глаз крупным планом; f — общий снимок фасеточного глаза, на котором стрелками отмечены отдельные омматидии; g — тот же вид, снятый в более контрастном свете (видны «прогалы» на месте утраченных омматидиев); h — отдельный омматидий, i — розетка из восьми ретинальных клеток; j — она же, снятая в другом освещении (красные стрелки показывают рабдом, на врезках продемонстрировано положение отдельных клеток и их схематичная зарисовка); k — вид сбоку на отделившийся от фасеточного глаза омматидий, видна роговичная линза и небольшой кристаллический конус (отмечен красной стрелкой). Иллюстрация из обсуждаемой статьи в Scientific Reports
Подобные «усовершенствования» были важны по той причине, что, хотя омматидии A. koninckii не были (как у S. reetae) упакованы в ярко выраженные «корзинки», между ними все же были достаточно толстые стенки из кутикулы. Из-за этого между роговичными линзами (которые имели диаметр всего около 30 мкм) оставались промежутки, и падающий на глаз свет использовался не полностью.
Зато омматидии A. koninckii были окружены пигментными клетками: авторы обнаружили там характерные следы пигмента, причем у трилобитов это удалось сделать впервые. Казалось бы, зачем пигмент, если есть кутикулярные стенки? По-видимому, стенки были полупрозрачными, что и вызвало необходимость в появлении пигмента, как у современного жаброногого ракообразного Artemia salina (рис. 3, j).

Рис. 3. а — два омматидия фасеточного глаза A. koninckii разной степени сохранности (отмечены красной и розовой стрелками); b — один из омматидиев в большем увеличении (видно, что рабдом окружен сферическими элементами, предположительно, представляющими собой палисад); c–h — еще несколько розеток ретинальных клеток с рабдомами, окруженными палисадом (особенно хорошо он виден на изображении f); i — зарисовка по фото с рис. 2, k (предполагаемый кристаллический конус окрашен синим); j — полупрозрачная голова жаброногого ракообразного Artemia salina, на снимке виден светоизолирующий пигмент внутри фасеточных глаз; k — вид на фасеточный глаз с другой стороны (стрелкой отмечен отделившийся омматидий с изображения i); l и m — реконструкция того, из какой части глаза и как омматидий мог отделиться (его траектория показана красной стрелкой); n — фасеточный глаз современного шершня обыкновенного (Vespa crabro) и его зарисовка, также показаны пучок ретинальных клеток с рабдомом в продольном разрезе (внизу справа) и поперечном (в виде розетки клеток). Условные обозначения: ru — рецепторная единица (отдельный омматидий), c — промежуток на месте утраченного омматидия, сс — кристаллический конус, p — палисад, pc — пигментные клетки, rc — рецепторные клетки, rh — рабдом, L — линза. Иллюстрация из обсуждаемой статьи в Scientific Reports
Один из омматидиев отделился от фасеточного глаза (рис. 3) и лежал сбоку от него. Особенной удачей было то, что он при этом повернулся на бок, — благодаря этому ученые смогли подробно изучить его строение в продольном разрезе. Оказалось, что он имеет крупную линзу, но зато маленький кристаллический конус толщиной всего около 5 мкм (у современных форм обычно наоборот, что отражено на рис. 1). Почему так? Похоже, основную «работу» по преломлению света в омматидиях A. koninckii брала на себя роговичная линза, которая в них, во-первых, была почти сферической формы, а во-вторых, по-видимому, имела в составе много кальцита, который обладает лучшими преломляющими свойствами, чем хитин. В кальцитовой природе роговичных линз трилобитов авторы обсуждаемой статьи, в отличие некоторых своих коллег (см. Палеоэнтомологи усомнились в уникальности глаз трилобитов, «Элементы», 13.09.2019), не сомневаются.
На том же рис. 3, i видно, что длина омматидия примерно равна его ширине. Это необычно: у современных членистоногих с фасеточными глазами омматидии имеют ярко выраженную вытянутую форму (опять же, см. рис. 1). Чем короче рабдом, тем меньше фотонов улавливают рабдомеры. При этом между рабдомерами не было зазоров (как в суперпозиционных фасеточных глазах), а значит у A. koninckii глаза были аппозиционными. Все указывает на то, что этот трилобит жил в хорошо освещенных приповерхностных водах и был активен днем.
При всем сказанном, глаза A. koninckii по своему устройству близки к глазам современных дневных насекомых (вроде шершня, см. рис. 3, n). Получается, что по сравнению с S. reetae, живший «всего лишь» примерно через 70 млн лет A. koninckii в этом плане сделал большой шаг вперед.
Описанная находка указывает на то, что фасеточные глаза возникли у членистоногих раньше и эволюционировали быстрее, чем считалось до сих пор. Впервые он появился, видимо, еще в докембрии — незадолго до того, как эволюционная линия трилобитов отделилась от линии предков насекомых и ракообразных (первые трилобиты, судя по ископаемым находкам, появились 540–520 млн назад, в начале кембрия). Она же говорит о том, что эволюция фасеточного глаза у трилобитов шла во многом в том же направлении, что и у этих более успешных в будущем групп.
Источник: elementy.ru





