













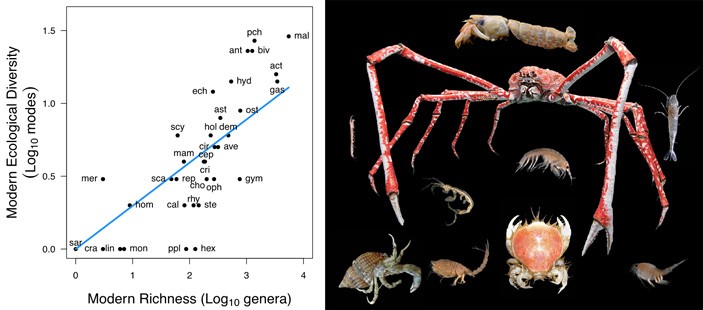


Рис. 1. У современных морских животных на уровне классов существует сильная положительная корреляция между экологическим и таксономическим разнообразием. По вертикальной оси — экологическое разнообразие (число разных образов жизни, характерных для представителей класса), по горизонтальной — таксономическое разнообразие (число родов). Обозначения классов: Act — Actinopterygii, Ant — Anthozoa, Ast — Asteroidea, Ave — Aves, Biv — Bivalvia, Cal — Calcarea, Cep — Cephalopoda, Cho — Chondrichthyes, Cir — Cirripedia, Cra — Craniata, Cri — Crinoidea, Dem — Demospongea, Ech — Echinoidea, Gas — Gastropoda, Gym — Gymnoleaemata, Hex — Hexactinellida, Hol — Holothuroidea, Hom — Homoscelomorpha, Hyd — Hydrozoa, Lin — Lingulata, Mal — Malacostraca, Mam — Mammalia, Mer — Merostomata, Mon — Monoplacophora, Oph — Ophiuroidea, Ost — Ostracoda, Pch — Polychaeta, Ppl — Polyplacophora, Rep — Reptilia, Rhy — Rhynchonellata, Sar — Sarcopterygii, Sca — Scaphopoda, Scy — Scyphozoa, Ste — Stenolaemata. Рисунок из дополнительных материалов к обсуждаемой статье в Science. Справа — представители класса Malacostraca (высшие раки), занимающего верхнюю правую позицию на графике. Изображение © Hans Hillewaert с сайта en.wikipedia.org.
Анализ данных по экологическому и таксономическому разнообразию современных и ископаемых групп морских животных показал, что связь между этими показателями не так проста, как считалось. Рост таксономического разнообразия (числа родов) не является ни прямым следствием, ни причиной высокого экологического разнообразия группы, измеряемого числом различных образов жизни. В последние 450 млн лет в морской фауне росла доля животных, относящихся к экологически разнообразным группам, а роль представителей экологически гомогенных групп снижалась. Но это происходило не из-за высокого темпа появления новых родов в экологически разнообразных группах, а из-за низкой скорости их вымирания. Иными словами, секрет успеха — не в высокой «рождаемости», а в низкой «смертности». По причинам, понятным лишь отчасти, «смертность» ниже в группах, освоивших большее число разных экологических ниш и образов жизни.
Если в каком-то классе животных или экосистеме много видов и родов (высокий уровень таксономического разнообразия), то логично предположить, что и экологическое разнообразие (число разных образов жизни, способов питания, передвижения и т. д.) в этом классе или в этой экосистеме тоже будет высоким. Данные по современной биосфере вроде бы подтверждают это предположение. Например, у современных морских животных на уровне типов, классов и отрядов прослеживается сильная положительная корреляция между таксономическим и экологическим разнообразием (рис. 1).
На первый взгляд этот факт может показаться банальным. Но в действительности вопрос о том, как формируется эта корреляция и как она менялась в ходе эволюции, изучен слабо. Теоретически здесь возможны разные механизмы и их комбинации. Например, можно предположить, что группы животных различаются по своей склонности к видообразованию: какие-то группы производят больше новых видов в единицу времени, какие-то меньше, просто в силу неких особенностей своего поведения, размножения и т. п. Чем больше появляется видов, тем острее межвидовая конкуренция за ресурсы, и тем чаще отбор поддерживает «расхождение признаков» и экологическую диверсификацию (как в известном случае с двумя видами вьюрков на острове Дафне, см.: Расшифрованы генетические основы быстрых эволюционных изменений размера клюва у дарвиновых вьюрков, «Элементы», 25.04.2016). В этом случае мы предполагаем, что экологическое разнообразие — это следствие, а таксономическое — причина.
Или, может быть, всё наоборот, и начинать рассуждение нужно с эволюционно-экологического потенциала группы: одни группы в силу своего плана строения, особенностей физиологии и прочего способны освоить много разных ниш, а другие сильнее ограничены в своем выборе. Тогда у первых будет больше возможностей не только для экологической, но и для таксономической диверсификации. Здесь мы предполагаем, что экологическое разнообразие — скорее причина, а таксономическое — в основном следствие.
Кроме того, нужно помнить, что таксономическое разнообразие зависит не только от скорости появления новых родов или видов, но и от скорости их вымирания. Может быть, экологически (или таксономически) разнообразные группы по какой-то причине имеют пониженный (или повышенный) темп вымирания?
Чтобы разобраться в этом, американские биологи собрали и проанализировали подробные данные по систематике и экологическому разнообразию современных и ископаемых морских животных. В базу данных вошли почти 20 000 родов, известных в ископаемом состоянии (как вымерших, так и доживших до современности), для которых моменты первого и последнего появления в палеонтологической летописи датированы с точностью до яруса (в палеонтологии это считается хорошим уровнем точности).
Для каждого рода был указан «образ жизни» (mode of life), который представляет собой комбинацию трех параметров: положение относительно дна (обитающий в толще воды; донный, но приподнятый надо дном, например, на стебельке; живущий на поверхности дна; частично зарывающийся и т. д. — всего шесть вариантов), подвижность (активно плавающий или ходящий, ползающий, передвигающийся лишь изредка, но не прикрепленный и т. д. — всего шесть вариантов) и способ питания (сестонофаг, соскребатель-обгрызатель, хищник и т. д. — тоже шесть вариантов). Всего, таким образом, получается 63 = 216 возможных вариантов, хотя далеко не все они используются реальными животными, а некоторые комбинации даже теоретически трудно себе представить (подробнее об этой классификации образов жизни рассказано в статье R. K. Bambach et al., 2007. Autecology and the filling of ecospace: Key metazoan radiations, откуда авторы ее и взяли).
Дополнительно использовались данные по современной морской фауне (всего 30 097 родов, включая роды, не известные в ископаемом состоянии).
У современных морских животных экологическое разнообразие (число разных образов жизни в отряде, классе или типе) положительно коррелирует с таксономическим разнообразием (числом родов в этом отряде, классе или типе), как показано на рис. 1.
Как выяснилось, в прошлые геологические эпохи такая корреляция тоже имела место. Однако она не всегда была такой же сильной, как сегодня. Наклон линии регрессии постепенно увеличивался (рис. 2). Иными словами, если в начале фанерозоя классы с большим числом родов лишь слегка превосходили по своему экологическому разнообразию классы с малым числом родов, то к концу фанерозоя разрыв сильно увеличился.

Рис. 2. Корреляция экологического и таксономического разнообразия в разные эпохи фанерозоя (на уровне классов). А — линии регрессии для разных отрезков фанерозоя. По горизонтальной оси — таксономическое разнообразие классов, по вертикальной — экологическое разнообразие. Цвета соответствуют разным временным отрезкам: современность (синий), кайнозой (желтый, 66,0–0,0117 млн лет назад), юра-мел (зеленый, 201,3–66,0 млн лет назад) и т. д. B — наклон линий регрессии для разных геологических ярусов. По вертикальной оси — наклон, по горизонтальной — время в млн лет назад. Вертикальными пунктирными линиями показаны пять великих вымираний фанерозоя, слева направо: позднеордовикское вымирание, позднедевонское, пермотриасовое, триасово-юрское, мел-палеогеновое. Рисунок из обсуждаемой статьи в Science
Рисунок 2, B показывает, что наклон графика зависимости экологического разнообразия от таксономического (величина, отложенная на графике по вертикальной оси) в течение фанерозоя рос не монотонно: он менялся скачками после массовых вымираний. Особенно хорошо заметны резкие подъемы после двух самых сильных вымираний, маркирующих границы эр: пермотриасового и мел-палеогенового.
По-видимому, это значит, что характерная для современной морской фауны сильная корреляция между экологическим и таксономическим разнообразием складывалась постепенно, причем массовые вымирания сыграли в этом не последнюю роль.
Как положено в подобных исследованиях, авторы проверили, не может ли обнаруженная тенденция быть артефактом, связанным с избирательной неполнотой палеонтологической летописи (в общем случае чем древнее эпоха, тем менее полны и точны палеонтологические данные), и пришли к выводу, что не может.
Затем исследователи посмотрели, как в разные эпохи соотносились скорости появления и вымирания родов в группах с высоким и низким родовым разнообразием. Понятно, что в более разнообразных группах разность скоростей появления и вымирания должна быть больше, чем в группах с низким родовым разнообразием. Однако повышенная разность скоростей может быть достигнута разными способами: например, увеличением скорости появления при неизменной скорости вымирания или, наоборот, снижением скорости вымирания при неизменной скорости появления (рис. 3, А).

Рис. 3. Соотношение скоростей появления родов (синии линии) и их вымирания (красные линии) в классах с низким и высоким родовым разнообразием (A, B: по горизонтальной оси — логарифм числа родов в классе) и в классах с низким и высоким экологическим разнообразием (C: по горизонтальной оси — логарифм числа образов жизни). Верхний ряд графиков (А) иллюстрирует общую идею о том, что высокий темп диверсификации (diversification), характерный для более таксономически разнообразных групп, может быть обусловлен разными причинами: повышенным темпом появления родов при неизменной скорости вымирания; повышенным темпом вымирания в сочетании с еще более высоким темпом появления и т. д. B, C — эмпирические данные для разных временных интервалов фанерозоя. Слева направо и сверху вниз: силур, девон, карбон, пермь, триас, юра, мел, даний (первый век палеогена), палеоген, плейстоцен. Рисунок из обсуждаемой статьи в Science
Палеонтологи давно заметили, что скорости появления и вымирания родов положительно коррелируют друг с другом: в группах с высоким темпом появления новых родов, как правило, повышена также и средняя скорость их вымирания. Поэтому иногда используется показатель «темп обновления родового состава» (volatility), который усредняет обе эти скорости и характеризует эволюционную динамику групп в целом, позволяя подразделить группы на «быстрые» и «медленные».
Выяснилось, что в кайнозое классы с высоким таксономическим разнообразием отличаются низкими темпами вымирания родов. Темп появления новых родов у них тоже ниже, чем в классах с низким родовым разнообразием. То есть в кайнозое в группах с высоким разнообразием новые роды появляются редко, а вымирают еще реже. Соответственно, общий темп смены родового состава у них низкий (ниже, чем в кайнозойских группах с низким родовым разнообразием). Проще говоря, в кайнозойских морях доминируют «медленные» группы. Типичные примеры доминирующих кайнозойских групп с очень низкими темпами появления и вымирания — это двустворчатые и брюхоногие моллюски. Средняя продолжительность существования родов в этих классах очень высока, смена родового состава происходит медленно.
В более древние времена, в мезозое и палеозое, ситуация была иной. Самые разнообразные классы отличались от классов с низким разнообразием не пониженными, а повышенными скоростями появления и вымирания. Роды в таких группах вымирали часто, но появлялись еще чаще. Иными словами, в палеозое и мезозое господствовали «быстрые» группы (рис. 3, B). Типичный пример очень разнообразной мезозойской группы с быстрой сменой родового состава — аммониты. Кстати, именно поэтому аммониты так важны для стратиграфии мезозоя: средняя продолжительность существования видов и родов аммонитов невелика, так что по найденным аммонитам, как правило, можно довольно точно определить возраст отложений. Брахиоподы и криноидеи (морские лилии) могут служить примерами «быстрых» групп, доминировавших в палеозойских донных сообществах.
Таким образом, в кайнозое по сравнению с предшествующими эрами изменился способ достижения классами животных высокого родового разнообразия. В палеозое и мезозое самыми таксономически разнообразными были классы, склонные к быстрой диверсификации («повышенной рождаемости»). В кайнозое на первый план вышли классы, представители которых наиболее устойчивы к вымиранию (классы с «пониженной смертностью»).
Связь между экологическим разнообразием и скоростями появления/вымирания отражена на рис. 3, C. На протяжении большей части фанерозоя экологически разнообразные классы были более «медленными», чем экологически однородные: у первых темп появления и вымирания родов был ниже, чем у вторых.
Анализ этих и других статистических закономерностей привел авторов к выводу, что экологическое разнообразие (то есть способность представителей группы осваивать разнообразные ниши) само по себе не является причиной ускоренной таксономической диверсификации. Скорее даже наоборот, ведь в экологически разнообразных группах темп появления новых родов в среднем ниже, чем в экологически однообразных. Поэтому нет оснований предполагать, что экологическое разнообразие группы (класса) непосредственно стимулирует появление новых родов. Данные не подтверждают и обратную идею — о том, что высокое таксономическое разнообразие стимулирует экологическую диверсификацию.
Связь между экологическим потенциалом группы и ее долгосрочным эволюционным успехом всё же существует, но она более тонкая и проявляется лишь на больших временных интервалах. Экологически разнообразные группы в среднем меньше страдали от массовых вымираний и быстрее восстанавливались после них по сравнению с экологически гомогенными группами. Поэтому в морской биоте поэтапно, от вымирания к вымиранию, росла доля животных, относящихся к экологически разнообразным классам.
Это проявляется, в частности, в том, что между экологическим разнообразием групп в палеозое и их таксономическим разнообразием в мезозое и кайнозое существует положительная корреляция. Иными словами, зная, насколько экологически разнообразным был данный класс в палеозое, мы можем предсказать, насколько таксономически разнообразным он будет в мезозое и кайнозое. В обратную сторону это не работает: зная таксономическое разнообразие групп в палеозое, нельзя предсказать их экологическое разнообразие в последующие эры.
В целом вырисовывается следующая картина. Существуют группы с быстрой сменой родового состава («быстрые» группы), которые обычно являются экологически гомогенными, и «медленные» экологически разнообразные группы. «Быстрые» группы иногда достигают очень высокого уровня таксономического разнообразия. Их слабость в том, что они плохо переносят сильные изменения среды. Поэтому во время массовых вымираний, когда происходят резкие скачки температуры, концентрации кислорода и pH морской воды, их таксономическое разнообразие сокращается очень резко, иногда вплоть до полного вымирания группы.
«Медленные» группы наращивают свое таксономическое разнообразие неторопливо, зато они спокойнее относятся к переменам условий. Это особенно ярко проявляется во время массовых вымираний: «медленные» группы переносят их с минимальными потерями. Поэтому доля животных, относящихся к «медленным» группам, в морской биоте поэтапно растет. Например, в палеозое доминирующими группами в донной фауне были трилобиты, брахиоподы, морские лилии и другие прикрепленные иглокожие — типичные «быстрые» группы с не очень высоким уровнем экологического разнообразия. Во время массового вымирания на рубеже перми и триаса эти группы пострадали очень сильно, и поэтому в мезозое баланс сместился в сторону преобладания «медленных» групп. После мел-палеогенового массового вымирания «медленные» группы, такие как брюхоногие и двустворчатые моллюски, игравшие в палеозойских сообществах второстепенную роль, окончательно заняли господствующее положение в донных сообществах (рис. 4).

Рис. 4. Слайды из моего курса лекций «История жизни на Земле», показывающие, как после массовых вымираний происходит смена доминирующих групп. По горизонтальной оси — время в млн лет назад. В обоих случаях более «быстрая», экологически однообразная группа (наружнораковинные головоногие, брахиоподы) во время крупных кризисов (пермотриасовое вымирание — около 250 млн лет назад, мел-палеогеновое — 66 млн лет назад) подвергалась более сильному вымиранию, после чего вперед выходила конкурирующая группа с более медленным темпом смены родового состава и более широким спектром доступных образов жизни (рыбы, двустворчатые моллюски)
Эти выводы не являются абсолютно новыми. Палеонтологи вообще-то давно заметили эти тенденции, в том числе избирательное вымирание экологически специализированных «быстрых» таксонов и постепенное накопление в биоте «медленных» эврибионтных (способных жить в широком спектре условий) групп (см., например: А. В. Марков, 2000. Возвращение черной королевы, или закон роста средней продолжительности существования родов в процессе эволюции). Однако новые данные делают эти выводы более обоснованными и детализированными.
Самый интересный вопрос, конечно, состоит в том, почему экологически разнообразные группы имеют в среднем более низкий темп смены родового состава и лучше переносят кризисы. Авторы попытались разобраться в этом, сопоставив вероятность вымирания родов с несколькими экологическими и физиологическими характеристиками, которые теоретически могут влиять на вероятность вымирания при изменении условий среды.
Анализ показал, что вероятность вымирания тесно связана, во-первых, с подвижностью. Активно передвигающиеся животные вымирают в среднем реже, чем неподвижные. Это логично, поскольку подвижность позволяет животным мигрировать и активно искать оптимальные для жизни места и ресурсы. Иными словами, подвижность должна повышать приспособляемость к переменам условий. Опять-таки, это не новый вывод: давно известно, что в течение фанерозоя отношение числа подвижных морских животных к неподвижным росло, причем не монотонно, а ступенчато, от вымирания к вымиранию (R. K. Bambach et al., 2002. Anatomical and ecological constraints on Phanerozoic animal diversity in the marine realm).
Во-вторых, вероятность вымирания у хищных животных ниже, чем у нехищных. Поэтому от кризиса к кризису растет доля хищников в общем разнообразии морских животных. В-третьих, «физиологически забуференные» животные вымирают реже, чем незабуференные. Под забуференностью понимается способность активно контролировать газообмен и другие физиологические процессы, о чем можно судить по степени развития дыхательной и кровеносной систем. Судя по палеонтологической летописи, «забуференные» животные легче переносят колебания концентрации кислорода, pH и температуры, и поэтому у них больше шансов пережить эпохи массовых вымираний. Эти закономерности тоже описаны в упомянутой выше статье Bambach et al., 2002.
В экологически разнообразных «медленных» классах действительно больше подвижных, хищных и «забуференных» форм, чем в экологически гомогенных «быстрых» классах. Именно этим во многом объясняется низкий темп вымирания родов в экологически разнообразных классах. Однако даже с поправкой на все эти три параметра представители экологически разнообразных классов всё равно имеют пониженную вероятность вымирания по сравнению с представителями экологически однородных классов. Значит, у первых есть еще какие-то преимущества, природу которых предстоит выяснить.
Таким образом, исследование, с одной стороны, просто подтвердило ряд давно замеченных закономерностей. С другой стороны, некоторые закономерности стали более понятными и обоснованными. Можно выделить два наиболее важных вывода с относительно высоким уровнем новизны:
1) Убедительно продемонстрирована отрицательная корреляция между экологическим разнообразием класса и темпом появления/вымирания родов. Чем шире спектр экологических ниш и образов жизни, потенциально доступных представителям класса, тем дольше существуют роды и тем медленнее обновляется их состав.
2) Те группы животных, которые доминируют в современных морях, добились своего господствующего положения не потому, что склонны к быстрой диверсификации, а потому, что их представители лучше переносят резкие изменения условий и благодаря этому реже вымирают.
Источник: elementy.ru





